

Мотивация и потребности личности
|
|
В психологии есть некая объединяющая позиция, согласно которой мотив это побудитель (сам термин происходит от лат. movere, что означает толкать, побуждать). Разногласия начинаются с психологического понимания самой функции побуждения. Кто-то считает мотив принадлежащим деятельности, а кто-то личности.
Мотив — это то, что отражается в сознании человека, побуждает его к деятельности, направляя ее на удовлетворение определенной потребности.
Потребность находит себя в конкретных мотивах, реализуется в них. Мотив — это ответ на вопрос: что нужно для удовлетворения потребности? Мотив принадлежит личности, которая “прикладывает” его к той или иной деятельности. Из личностного он становится мотивом деятельности как бы вторично.
В основе любой деятельности человека лежит мотив, побуждающий его к этой деятельности. Однако соотношение деятельности и мотива не однозначно. Тот или иной мотив, возникший у личности, не всегда в этой деятельности исчерпывается. В процессе деятельности мотив может измениться, и, напротив, при сохранности мотива может измениться выполняемая деятельность. Иногда формирование мотива опережает формирование деятельности, а иногда — напротив — отстает — и то и другое сказывается на ее результате.
Вопрос о том, откуда берутся мотивы, как они возникают, является в психологии личности одним из центральных. Та же самая концепция А. Маслоу не может ответить на этот вопрос: как известно, он построил “пирамиду” потребностей. Которые являются основой мотивов, — биологические потребности, потребность в безопасности (как нужда в порядке, устойчивости), потребность в уважении, престиже, и, наконец, потребность в самовыражении, в самоактуализации. Но Маслоу берет потребности индивида, “вырывая” его из системы общественных отношений, рассматривая их вне связи с обществом.
На данном этапе в психологии нет четкой и общепринятой классификации мотивов и потребностей. А. Н. Леонтьев образно отмечал, что понятие мотива превратилось для психологии в большой мешок, в котором сложены самые разнообразные вещи.
Мотивы различают
- Мотивы различают по виду тех потребностей, которым они отвечают. Поэтому возможны, например, мотивы высшие и низшие, материальные и духовные. Мотивы можно разложить на виды по любой из существующих классификаций потребностей, в частности по “пирамиде” А. Маслоу.
- Мотивы различают по форме отражения предмета потребности. Мотивом выступают самые разные психологические образования: нравственное понятие или представление, идеал, ценностная ориентация.
- По степени обобщения предмета потребности мотивы могут быть широкими и узкими, по времени существования и действия — кратковременными и долговременными. Широкие мотивы выполняют смыслообразующую функцию, тогда как узкие мотивы являются реальными побудителями. Например, человек длительное время выполняет какую-то неинтересную работу. Здесь узкими и кратковременными побудителями выступают прозаические мотивы: возможность поощрения, угроза наказания, просто необходимость действовать. Но человеку нужны какие-то более долговременные и широкие мотивы: мечты, будущая жизнь, нравственные ценности.
- Мотивы подразделяют по степени участия в них сознания. Потому мотивы могут быть осознанными и неосознанными, понимаемыми и реально действенными. Допустим, студент первокурсник знает, что учиться нужно для своего светлого будущего, для глубоких знаний. Но такой мотив вряд ли является для него реально действующим. Действительное побуждение к учению идет за счет исполнения заданий, подражания однокурсникам или значимым людям.
Установки
Среди неосознаваемых побуждений личности лучше всего изучены установки. Установкой в психологии обозначается неосознаваемое личностью состояние готовности, предрасположенности к деятельности, с помощью которой может быть удовлетворена та или иная потребность.
Установка — это готовность, предрасположенность определенным образом воспринять, понять, осмыслить объект или действовать с ним в соответствии с прошлым опытом.
Эксперименты Д. Н. Узнадзе и его сотрудников показали процесс формирования фиксированных установок, определяющих поведение личности. Предвзятость, составляющая сущность многих установок, либо является результатом поспешных и недостаточно обоснованных выводов из истории личного опыта человека, либо это результат некритического усвоения стереотипов мышления — стандартизированных суждений, принятых в определенной общественной группе.
Установки по отношению к различным фактам общественной жизни могут быть позитивными и негативными. Психологическими исследованиями в структуре установки выделены три составляющие. Когнитивная подструктура есть образ того, что готов познать и воспринять человек; эмоционально-оценочная подструктура есть комплекс симпатий и антипатий к объекту установки; поведенческая подструктура — готовность определенным образом действовать в отношении объекта установки.
К неосознаваемым мотивам также относят влечения, которые определяются как неопредмеченное побуждение.
Как отмечал А. Н. Леонтьев, у человека в голове нет мотива. Он представлен в сознании опосредованно, в виде смысла. Реальным побудителем, задающим результативность деятельности, является мотив — неосознанный, более прагматичный.
Сказанное выше позволяет заключить, что мотив является не просто одной из составляющих деятельности, а выступает в качестве компонента сложной системы — мотивационной сферы личности. Под мотивационной сферой личности понимается вся совокупность ее мотивов, которые формируются и развиваются в течение жизни. В целом эта сфера подвижна и развивается в зависимости от обстоятельств. Но некоторые мотивы относительно устойчивы и, доминируя, образуют как бы стержень всей сферы (в них проявляется направленность личности).
Эмоции и потребности
Взаимосвязь эмоций и потребностей отмечается практически всеми исследователями. В простейших случаях констатируется зависимость эмоций от удовлетворения или неудовлетворения потребностей, в первую очередь органических. В более сложных формах уже говорят не о зависимости эмоций от потребностей, а о взаимосвязи эмоций и потребностей. Так, Рубинштейн отмечает, что:
«взаимоотношение эмоций с потребностями может проявляться двояко — в соответствии с двойственностью самой потребности, которая, будучи испытываемой индивидом нуждой его в чем-то ему противостоящем, означает одновременно и зависимость его от чего-то и стремление к нему».
Эмоция в этом случае отражает и удовлетворение неудовлетворение потребности, с одной стороны, и стремление, влечение к тому, что заключает в себе опредмеченную потребность, с другой.
Исходя из общего положения, высказанного выше, можно утверждать, что развитие эмоций и чувств отражает как развитие потребностей индивида, так и сами эмоции, которые, выступая в роли источника стремлений и влечений, начинают выражать активную сторону потребности и ведут к многообразию предметной реализации конкретной потребности.
Развитие самих потребностей и многообразия предметной реализации конкретных потребностей ведет к развитию эмоций и чувств. Окружающий мир становится для субъекта чувственно значимым, а восприятие им такого мира сопровождается переживаниями. Человек с богатыми потребностями и широкой опредмеченностью этих потребностей живет богатой эмоциональной (чувственной) жизнью.
Таким образом, с одной стороны, потребности являются условием возникновения эмоций и чувств, а с другой стороны, сами эмоции и чувства могут выступать в роли потребностей и мотивов, определяющих поведение человека.
§11. Биологические мотивации
как внутренние детерминанты
поведения
Принимая в качестве необходимого фактора целенаправленного поведения наличие биологической потребности, И. П. Павлов указывал, что нелепо было бы отрицать субъективный мир, как и психологию — науку, изучающую явления этого субъективного мира. Проблема ставилась так: каковы те внутренние состояния животных, которые придают их поведению целенаправленный характер. Некоторые ученые полагали, что желания, побуждения, предшествующие действию и определяющие его, составляют движущие силы или мотивы действия. Термин мотивации буквально означает то, что вызывает движение. Этот термин долгое время не использовался в физиологии и входил исключительно в лексикон психологов.
Мы уже отмечали (см. гл. 2), что первоначальный смысл термина инстинкт — это побуждение к действию. Еще И. М. Сеченов полагал, что животное стремится удержать, продлить приятное чувственное возбуждение и, наоборот, избавиться от действия раздражений, сопровождающихся неприятными ощущениями. Таким
образом, мотивации оказывались поставленными в прямую и двустороннюю взаимозависимость от эмоций. И. М. Сеченов ввел категорию «желаний», куда он относил голод, жажду, половое чувство. На этих желаниях и строятся «страстные психические рефлексы», по Сеченову, которые сопровождаются томительными отрицательными переживаниями, а сами действия, т. с. удовлетворение страстного желания, имеют положительный характер.
«Жизненные потребности, - писал И. М. Сеченов,— родят хотения и уже эти ведут за собой действия; хотение тогда будет мотивом или целью, а движение — действием или средством достижения цели... Без хотения как мотива или импульса движение было бы вообще бессмысленно»'.
И. П. Павлов указывал на значение основных влечений — голода, жажды, полового чувства п возникновении «рефлекса цели». Вся жизнь, вся ее культура делается рефлексом цели, который является важнейшим фактором жизни, начиная с капитальнейшей области — воспитания.
Этологи назвали целенаправленное поведение аппетентным, куда включались все случаи поискового поведения пищи, полового партнера, материала для постройки гнезда и т. д. В изучении эндогенных пусковых механизмов поведения большая роль отводилась гормональным системам. Нще в 1923 г. А. А. Ухтомский подчеркивал роль гормональных механизмов в первичном возникновении некоторых доминантных состояний, которые в дальнейшем могут воспроизводиться и модулироваться условно-рефлекторными механизмами.
Физиологические потребности. Известно, что нормальный уровень обмена веществ и энергии может поддерживаться только при определенном содержании в организме ряда продуктов жизнедеятельности, при их постоянном потреблении, накоплении и разрушении. Увеличение или снижение содержания этих продуктов в организме существен-
1 Сеченов И. М. Изб. произв. М., АН СССР, 1952. Т. I. С. 516.
но меняет его жизнеспособность. Приобретенные в процессе эволюции механизмы саморегуляции обеспечивают организму поддержание концентрации этих веществ во внутренней среде на относительно постоянном уровне (гоме-остаз). Этот уровень основных жизненно важных веществ в организме характеризуется как определенные константы внутренней среды. К таковым относятся уровень содержания в крови питательных веществ, ее газового состава, осмотического давления, температуры и проч.
В результате непрерывного обмена веществ внутренние константы крови имеют тенденцию к колебаниям (гомео-кинез), но восстанавливаются до требуемого уровня задолго до наступления существенных сдвигов в обмене веществ. Однако такой автоматический саморегулирующийся механизм (с внутренним контуром обратной связи) действует б определенных пределах и с разной скоростью. Но в тех случаях, когда отклонение от нормальных констант становится слишком значительным и не поддается коррекции с помощью' автоматической саморегуляции, вступают в действие более высокие уровни, связанные с потреблением определенных веществ из внешней среды или выведением продуктов обмена из организма. Эти более высокие уровни саморегуляции (с внешним контуром обратной связи) достигаются благодаря целенаправленному поведению.
Любой живой организм может существовать лишь при периодическом потреблении из внешней среды питательных веществ, воды, минеральных солей н т. д. Такие состояния получили название первичных физиологических потребностей. Однако термин потребность может трактоваться и более широко. Исходя из представлений В. И. Вернадского и А. А. Ухтомского о присущей живой системе тенденции самосохранения и саморазвития, П. В. Симонов (1987) предлагает
под потребностью рассматривать специфическую (сущностную) силу живых организмов, обеспечивающую их связь с внешней средой для самосохранения и саморазвития, как источник активности живых систем к окружающем мире.
П. К, Ано:.ин (1968) и К- В. Судаков (1971, 1986) в исследовании проблемы потребности исходят из общебиологического принципа выживания как конечной цели лч/Гюго поведения животных.
Истинная метаболическая потребность возникает в тканях организма, однако сигнализация о ней генерируется прежде всего в специальных высокочувствительных рецепторах. Восприятие потребности происходит нервным и гуморальным путем, а соответствующая нервная сигнализация адресуется в специальные подкорковые отделы мозга и прежде всего в ядра гипоталамуса — важнейшего отдела промежуточного мозга (рис. 19). Возбуждение этих и связанных с ними центров и формируют состояние мотивации. Приведем два основных определения этого термина.
Мотивация представляет собой возникающие под влиянием первичных изменений во внутренней среде эмоционально окрашенные состояния организма, характеризующиеся избирательными активирующими влияниями специальных подкорковых аппаратов на кору головного мозга и дру-1 ие его отделы и направляющая поведение животного на удовлетворение исходной потребности
(К. В. Судаков, 1971). Соответственно потребностно-информационный подход к пониманию деятельности мозга позволяет считать, что
мотивация есть физиологический механизм акти-вировання хранящихся в памяти следов (эн-грамм) тех внешних объектов, которые способны удовлетворить имеющуюся у организма потребность, и тех действий, которые способны привести к ее удовлетворению (П. В. Симонов, 1987).
Если первое определение мотиваций в большей мере адресуется к первичным физиологическим потребностям, то второе объясняет целенаправленный характер поведения во всем разнообразии его психофизиологических проявлений.
Виды мотиваций. Если исходить из определения поведения как формы жизнедеятельности, которая меняет вероятность и продолжительность контакта с объектом удовлетворения имеющейся у организма потребности (П, В. Симонов, 1979), то вся наша жизнь представляется как непрерывная цепь фор-
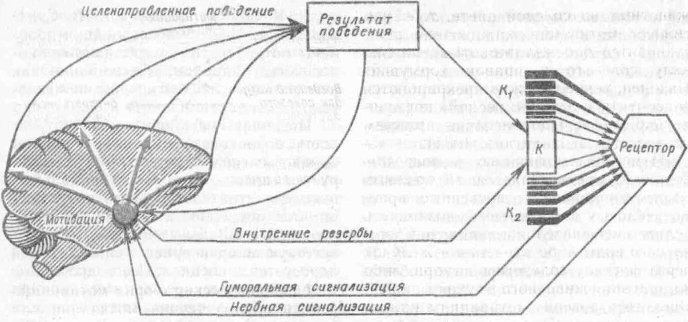
Рис. 19. Принципиальная схема сяморегу-ляторных процессов, порождающих в центральной нервной системе мотивационное побуждение (по К. В. Судякову, 1972):
К — константа крови, отклонение константы от жизненно важного уровня (Ki и К2) череч возбуждение рецепторов или гуморальным путем вызывает в центральной нервной системе мотиваиионное возбуждение. Последнее приводит к целенаправленному повелению и удовлетворению исходной потребности организма
мирующихся целей и их достижении или неудач.
Превращение актуализированной потребности из побуждения в цель и составляет содержанке процесса мотивации целенаправленного поведения.
Наиболее распространенная классификация видов мотиваций предложена Н. Миллером (1960). Он выделяет низшие, или первичные мотивации, называя их также инстинктивными или висцеральными. Эти же состояния И. П. Павлов рассматривал как основные влечения и относил к ним голод, жажду, страх, половое чувство. К ним же могут быть отнесены и наиболее примитивные состояния, которые побуждают к актам мочеиспускания, дефекации и т. д.
Вторую группу мотиваций составляют высшие или вторичные мотивации, которые приобретаются в течение индивидуальной жизни и, хотя и строятся на основе генетически заданных потребностей, в значительной мере опираются на накопленный индивидуальный опыт. Сюда могут быть отнесены все виды мотиваций, которые возникают по законам условного рефлекса.
Методы изучения биологических мотиваций. Одним из основных внешних проявлений мотиваций является целе-
направленное поведение. Предложенная П. К. Анохиным (1932) методика свободного выбора пищи животным оказалась весьма демонстративной, ибо предпочтение животным определенного вида пищи (мясо, молоко, солевой раствор и пр.) свидетельствовало о наличии соответствующей исходной мотивации.
Условно-рефлекторная методика позволяет придать различным внешним агентам функции сигналов получения определенных видов пищи или воды и тем самым определить степень соответствия внешней сигнализации естественно или искусственно сформированной внутренней мотивации. Например, у собаки вырабатывают условные рефлексы в форме побежки на сигнал стука метронома 60 ударов в минуту к левой кормушке, в которой всегда подается пресная пища, а на сигнал 120 ударов в минуту — к правой кормушке, побежка к которой подкрепляется пи-шей разной степени солености. Определив предельную степень солености пиши, которую поедает животное (солевой порог), собаку переводят на искусственную солевую диету с одновременным ограничением воды. Естественно, что чем длительнее содержание
животных на солевой диете, т. е. чем сильнее мотивация жажды, тем реже становятся побежки животных по сигналу соленого к правой кормушке. Наконец, такие побежки прекращаются вовсе, хотя к левой «пресной» кормушке побежки сохраняются на прежнем уровне (И. П. Никитина, 1985).
Измерить мотивацию непосредственно не представляется возможным, поэтому используют косвенные приемы. Мотивация может возникнуть вследствие длительного лишения животного воды либо введения в желудок через фистулу солевого раствора, либо кормления животного сухой пищей. Оценить уровень мотивации жажды можно несколькими способами (рис. 20): путем измерения общего объема выпитой животным воды, частоты нажатия на рычаг для получения питьевого подкрепления или предельного количества хинина, добавленного в воду, при котором животное еще продолжает ее пить.
Электрическое раздражение определенных нервных центров, расположенных в гигюталамической области, сопровождается не только возникновением первичных мотиваций, но и соответствующим поведением, ведущим к их удовлетворению. Так, например, у крыс предварительно вырабатывают условный рефлекс нажатия на рычаг, за которым следует пищевое подкрепление. При раздражении латеральной ги-поталамической области даже накормленные крысы снова начинают нажимать на рычаг (Н. Миллер, 1960).
Особый интерес для физиологии мотиваций представляют эксперименты с самораздражением (Олдс, 1958). Оказалось, что раздражение определенных структур головного мозга (прежде всего ядер гипоталамуса) через вживленные электроды приводит к тому, что животное начинает нажатием на рычаг само включать ток, чтобы наносить себе раздражения. Эти мотивации обладают значительной энергетической силой, и животные готовы преодолевать сложные препятствия ради достижения эффекта самораздражения.
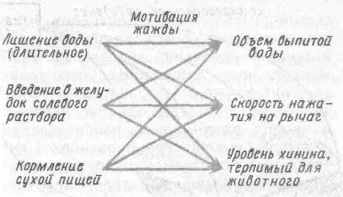
Рис. 20. Возможные способы изображения взаимосвязи между тремя независимыми переменными, которые позволяют измерять потребление воды (но Э. Мэннингу, 1982)
Физиологические теории мотиваций.
Первая группа теорий мотивации основана на представлении о сигнализации от периферических органов тела. В. Кеннон 1932, 1934), Д. Хебб (1949) полагали, что мотивации определяются стремлением организма избежать неприятных эмоциональных ощущений, сопровождающих различные побуждения. Животное утоляет жажду, чтобы избавиться от неприятной сухости в полости рта и глотки, поедает пищу, чтобы избавиться от мышечных сокращений пустого желудка, и т. д.
Дальнейшие поиски привели к созданию другой группы теорий, в которых основное внимание уделялось гуморальным факторам мотиваций. Так, голод связывался с возникновением так называемой голодной крови, т. е. крови с существенным отклонением от обычной разницы в концентрации глюкозы в артериальной и венозной крови. Это и рассматривалось в качестве первопричины «голодных» сокращений желудка. Мотивация жажды также оценивалась как следствие изменения осмотического давления плазмы крови или снижения внеклеточной воды в тканях, что может оцениваться специальными рецепторами (осмо- и волюмрецепторы) . Половое влечение ставилось в прямую зависимость от уровня половых гормонов в крови.
Для оценки справедливости гуморальных теорий мотиваций существенный интерес представляют наблюдения за двумя парами сросшихся близнецов-сакропагах, выполненные Т. Т. Алек-
сеевой (1958). У этих близнецов была общая система кровообращения и раздельная центральная нервная система. Верхняя часть пищеварительного тракта у них также была раздельной. Оказалось, что кормление одной из девочек, а значит равномерное распределение питательных веществ в кровеносной системе обоих близнецов, никогда не приводило к насыщению второй девочки. Наоборот, у нее длительное время могла поддерживаться потребность в пище. Эти наблюдения, а также многочисленные наблюдения на животных показали, что гуморальные факторы не могут быть первопричиной возникновения биологических мотиваций.
По-видимому, как возникновение мотиваций, так и их удовлетворение имеет множественный генез, в основе которого лежит нейрогуморальный механизм периферического и центрального уровней.
В самих мозговых структурах обнаружены чувствительные нервные клетки, специализированные на восприятии колебаний в содержании определенных химических веществ, а также физических параметров питающих их жидкостей. Основным аппаратом, содержащим такие центральные рецепторы, является гипоталамус. И. Стелл ар (1954), выдвинув гипоталамическую теорию мотиваций, полагал, что гипоталамус и является вместилищем «центрального мотивационного состояния».
Рассмотрим для примера некоторые гипоталамические механизмы жажды. Во-первых, в латеральных ядрах гипоталамуса обнаружены осморецепторы, детектирующие осмоляльность циркулирующей крови. Во-вторых, введение в латеральный гипоталамус солевого гипертонического раствора вызывает усиление потребления воды сверх физиологических потребностей (похожую реакцию вызывает и электрическое раздражение этих же структур). В-третьих, повреждение латерального гипоталамуса нарушает регуляцию водного баланса, а повреждение переднего гипоталамуса и преоптического ядра полностью устраняет способность к потреблению воды, не лишая, однако, животных воз-
можности принимать пищу. И наконец, в-четвертых, раздражение «питьевой» зоны гипоталамуса провоцирует целый комплекс поведенческих реакций, связанных с поисками воды.
Хорошо известно, что прекращение питья наступает задолго до того, как восстановятся физиологические константы крови. Показано также, что ни растяжение желудка, ни всасывание воды не имеет решающего значения в регуляции потребления воды. По этому поводу высказывается наиболее вероятное предположение — прекращение приема воды наступает в результате обратной связи от рецепторов ротовой полости, с помощью которой отмеряется количество поглощенной воды, и через амигдалу тормозится реакция питья (Н. Мильнер, 1973).
Теперь рассмотрим некоторые теории мотивации голода, которые связывают это состояние опять же со структурами гипоталамуса. Наиболее последовательные поиски ведутся Б. К. Анандом и Д. Р. Бробеком (1951), которые считают, что в латеральном гипоталамусе располагается центр питания, а в медиальном гипоталамусе — центр насыщения. Авторы допускают, что центры питания и насыщения находятся в реципрокных отношениях. Потребление пищи определяется тормозными влияниями латеральной области на медиальную.
С. П. Гроссман (1960—1969) вводил в гипоталамус накормленных крыс биологически активные вещества. Введение в латеральный гипоталамус адренергических веществ (в отличие от холинергических) вызывало усиленное потребление пищи. Введение же холинергических веществ провоцировало питьевые реакции. При этом возникал целый комплекс ранее заученных водо-и пищедобывательных двигательных реакций.
Сегодня является бесспорным положение о том, что
гипоталамические структуры не мoгyт paссматриваться в качестве единственных аппаратов пищевого поведения.
Лимбический уровень (включая гипоталамические и ретикулярные механизмы) обеспечивает лишь примитивно
организованные лишенные выраженной целенаправленности типы пищевого поведения, так называемые пищевые автоматизмы. Направленный же поиск пищи, ее оценка и выбор, а также формирование избирательного аппетита требуют участия коры больших полушарий головного мозга. Следовательно, только объединенное участие гипоталамуса, лимбических образований и коры головного мозга формирует целостное пищевое поведение животных.
При этом исключительная роль гипоталамуса не вызывает никаких сомнений:
этот отдел мозга как комплексный детектор физиологических дисбалансов (температуры, питательных веществ, воды, гормонов, минеральных солей) играет доминирующую роль в отношении путей использования ресурсов организма.
Практически ни одну функцию организма нельзя себе представить вне прямой или косвенной зависимости от гипоталамуса.
Работами К. В. Судакова (1971, 1986) было показано, что каждое мотива ционное возбуждение строится на основе специфических химических механизмов, активирующих влияния подкорковых мотивационных центров на кору головного мозга. Введением центральных адренолитиков (аминазина) удается избирательно заблокировать активацию коры при болевом раздражении. Активация же коры головного мозга при пищевом возбуждении голодных животных избирательно блокируется холино-литическими веществами.
Наряду с вышесказанным существует и крайняя точка зрения П. Мак-Лина (1970) о жесткой привязке мотиваций к аппаратам лимбической системы. Он считает, что в гипоталамусе локализованы эмоционально окрашенные поведенческие реакции, связанные с рефлексами избегания, нападения, поиска, добывания пищи, воды и пр. В амигдале сосредоточены мотивации голода, жажды, защиты от внешних вредных влияний. Перегородка связана с половым поведением. Противоположной позиции, которая кажется нам более справедливой, придерживается Р. Л. Аисааксон (1978), который отмечает, что не существует
одиночных анатомических образований для одиночных поведенческих функций, как не существует единых поведенческих функций, которые могли бы быть приурочены к анатомическим образованиям таким образом, чтобы эти функции оказывались полезными в любых обстоятельствах и при всех внешних условиях.
Доминирующее мотивационное возбуждение. В каждый момент времени доминирует та мотивация, в основе которой лежит наиболее важная биологическая потребность. Сила потребности, т. е. величины отклонения физиологических констант или концентрации соответствующих гормональных агентов, получает свое отражение в величине мотива ционного возбуждения структур лимбической системы и определяет его доминантный характер. Консервативный характер доминанты заключается в ее инертности, устойчивости, длительности. В этом ее большой биологический смысл для организма, который стремится к удовлетворению биологической потребности в случайно и постоянно меняющейся внешней среде. В физиологическом смысле такое состояние доминанты характеризуются определенным уровнем возбудимости центральных структур, обеспечивающей их высокую отзывчивость и «впечатлительность» к разнообразным воздействиям. Но однажды проявленная доминирующая мотивация возобновляется и живет даже в порядке чисто нервно-рефлекторного фактора. Поэтому наряду с консервативным началом в доминирующей мотивации как векторе поведения содержится и динамическое начало, поскольку из множества новых подкрепляющих впечатлений происходит непрерывный подбор «пригодного», нужного, имеющего непосредственную связь.
Первичный очаг возбуждения при формировании доминирующих биологических мотиваций (голод, жажда, секс) возникает в гипоталамических образованиях мозга (Р. А. Павлыгина, 1956). Эти первичные очаги, в свою очередь, ведут к появлению вторичных очагов в других отделах мозга, где
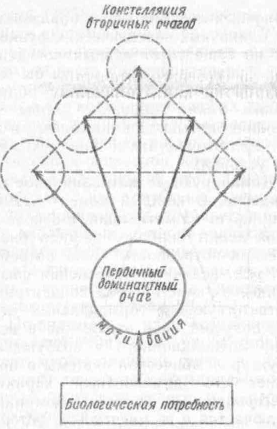
Рис. 21. Схема соотношения первичных и вторичных очагов при формировании доминирующей констелляции моягопых образований (по Л. С. Батуеву, 1986)
имеются предпосылки для продолжительного удержания следовых процессов. Кора головного мозга с ее непрерывной и разномодальной активацией располагает всеми предпосылками для создания в ней вторичных очагов стационарного возбуждения. Принципиальным отличием таких очагов от первичных является исходная причина их возникновения. Так, если первичный очаг в гипоталамусе возникает и поддерживается в первую очередь за счет гуморальных факторов, то вторичные очаги в коре больших полушарий являются проекционными по своему происхождению и поддерживаются за счет широкой конвергенции к ним множества суммирующихся здесь сенсорных влияний.
Рассмотрим несколько конкретных примеров. Содержание животных на длительной солевой диете, формирующее у них мотивацию жажды, создает в латеральном гипоталамусе очаг повышенной возбудимости к вкусовым и внсцерохи ми чески м раздражениям
(А. С. Батуев, Б. Г. Гафуров, 1988). Снимается же подобный очаг введением в организм дополнительных количеств жидкости.
Ф Итак, мотивационное возбуждение, обусловленное преобладающей потребностью организма, формирует в гипоталамусе первичный очаг повышенной возбудимости, обладающий свойствами доминантного очага. Благодаря этому доминантному очагу быстро образуются условно-рефлекторные связи с теми или иными фа.кторами окружающей среды (вторичные очаги), которые по своей функциональной значимости могут быть сравнимы с эндогенными механизмами самого мотива-ционного состояния (рис. 21).
Нейрофизиология мотиваций. К. В. Судаков (1986) сформулировал основные положения о принципах нейрофи-зиологического обеспечения доминирующих мотиваций: 1. Любая биологическая мотивация обусловлена соответствующей метаболической потребностью и носит системный характер.
2. Потребность трансформируется ней-рогуморальным путем в возбуждение гипоталамических центров, которые активируют другие структуры мозга, в том числе и кору больших полушарий.
3. Корковые и лимбические структуры мозга оказывают специфические для каждой мотивации нисходящие возбуждающие и тормозные влияния на гипоталамические мотивационные центры. 4. Каждое мотивационное возбуждение представляет собой специфическую клеточную и молекулярную интеграцию корково-подкорковых структур. Избирательное возбуждение одних структурных образований сочетается с избирательным торможением других.
Мотивационные возбуждения существенно меняют конвергентные и дискриминационные способности нейронов разных отделов мозга. Например, раздражение пищевого центра латерального гипоталамуса приводило к тому, что нейроны сенсомоторной коры, ранее не отвечавшие на световые,
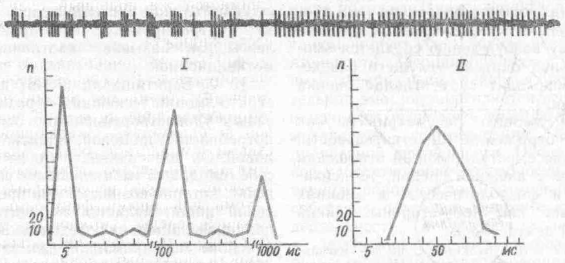
Рис. 22. Типичный характер нейронной активности у голодного (/) и накормленного кролика (//) (по К. В. Судакову, 1986):
под нейронограммой — гистограммы распределения межимнульсных интервалов
звуковые и гуморальные раздражения, приобрели способность реагировать на них. С другой стороны, нейроны коры, не отвечавшие ранее на введение кроликам морковного сока, начинали реагировать на эти раздражения после стимуляции центра «голода» латерального гипоталамуса. Создается впечатление, что мотивационное возбуждение «настраивает» нейроны различных областей мозга на подкрепляющие воздействия.
Мотивационное возбуждение находит свое выражение в специфическом распределении межимпульсных интервалов и образовании характерной пачечной активности (рис. 22). Эта активность усиливается при возрастании потребности, при возникновении препятствий к удовлетворению доминирующей мотивации. Достижение же цели поведенческого акта сменяется в нейронограмме регулярной активностью.
Возбуждения, поступающие в мозг при подкреплении, адресуются к различным нейронам и оставляют «след» в их деятельности, формируя сложную нейродинамическую констелляцию. С ней и происходит сравнение параметров достигнутых результатов.
В. Г. Зилов (1978) показал, что в формировании различных биологи-
ческих мотивации участвуют одни и те же нейромедиаторы, однако в различных комбинациях в разных структурах мозга, что свидетельствует о специфической нейрохимической интеграции конкретного мотивапионного возбуждения. Последнее проявляется также и в общей чувствительности отдельных нейронов к нейромедиаторам и оли-гопептидам (К. В. Судаков, 1986). Способность ряда пептидов (ангиотензин И, бета-липопротеин, пентогастрин и др.) при внутрижелудочковом введении избирательно продуцировать или подавлять определенные биологические мотивации открывает путь к изучению молекулярной интеграции мотива-ционного возбуждения.