

Половое размножение
 |
 |
|
Генеративные органы растений: архегоний (вверху) и антеридий (внизу)
|
Основу полового размножения (или воспроизведения) составляет половой процесс — слияние мужской и женской гамет. Органы, образующие их, — антеридии и архегонии — развиваются на растениях полового поколения — гаметофитах. У водорослей (за исключением харовых) они одноклеточные, у высших растений — всегда многоклеточные, как и спорангии.
Внутренние фертильные клетки антеридия составляют сперматогенную ткань, образующую мужские гаметы (сперматозоиды). Типичный зрелый архегоний состоит из расширенногобрюшка и узкой шейки, заполненной шейковыми канальцевыми клетками. В процессе эволюции количество фертильных клеток сократилось до единственной яйцеклетки. Оплодотворение внутреннее, в брюшке архегония. Образующаяся при этом зигота представляют собой первую клетку спорофита.
Выше дана самая общая характеристика полового размножения растений; вместе с тем у отдельных групп растений оно имеет свои существенные особенности.
У покрытосеменных растений органом размножения является цветок. Для выяснения функционирования цветка необходимо проследить, какие процессы происходят в тычинках и пестиках. Как сказано выше, тычинка состоит из тычиночной нити и пыльника. Каждый пыльник образован двумя половинками, в которых развивается по две пыльцевые камеры (гнезда) — микроспорангии. В гнездах молодого пыльника имеются особые диплоидные клетки —микроспороциты, или материнские клетки микроспор. Каждый микроспороцит претерпевает мейоз и образует четыре микроспоры. Здесь же, внутри пыльцевого гнезда, микроспора увеличивается в размерах, ядро ее делится митотически, и образуется два ядра — вегетативное и генеративное. На поверхности бывшей микроспоры образуется прочная целлюлозная оболочка с несколькими округлыми порами, сквозь которые в конечном итоге прорастают пыльцевые трубки. В результате этих процессов каждая микроспора превращается в пыльцевое зерно (пыльцу) —-мужской гаметофит цветковых растений (рис. 8.18).
У однодольных растений в пыльцевом зерне, находящемся в пыльнике, генеративное ядро делится митотически с последующим образованием двух неподвижных мужских гамет — спер-миев. У двудольных образование спермиев происходит позже, когда пыльца попадает на рыльце пестика. Таким образом, зрелое пыльцевое зерно состоит из двух (вегетативной и генеративной) или из трех (вегетативной и двух спермиев) клеток.
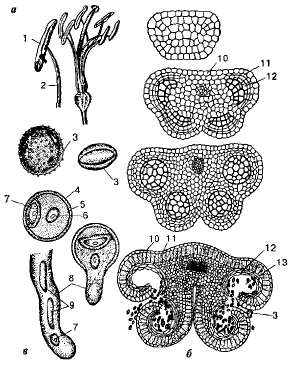
Рис. 8.18. Тычинки: а — общий вид тычинок; б —развитие пыльцевых гнезд; в —- пыльца и ее прорастание; I — пыльник; 2 — тычиночная нить; 3 — пыльца; 4 —экзина; 5 — интина; б — генеративное ядро; 7 — вегетативное ядро; 8 — пыльцевая трубка.; 9 — два спермия; 10 — эпидермис; И — фиброзный слой; 12 —спорогенная ткань; 13 — гнездо пыльника.
Образование женского гаметофита происходит в семязачатке (семяпочке), находящемся внутри завязи пестика (рис. 8.19). Семязачаток — это видоизмененный мегаспорангий (нуцеллус), защищенный покровами. Покровы на верхушке не срастаются и образуют узкий канал — пыльцевход. В нуцеллусе, вблизи пыльцевхода, начинает развиваться диплоидная клетка — мегаспороцит (макроспороцит). Он делится мейотически, давая четыре гаплоидные макро- или мегаспоры, обычно расположенные линейно. Три мегаспоры вскоре разрушаются, а четвертая, наиболее удаленная от пыльцевхода, развивается в зародышевый мешок.
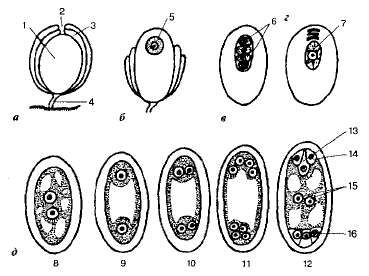
Рис. 8.19. Образование макроспор в семязачатке (а—в) и развитие зародышевого мешка (г—д): 1 —нуцеллус; 2 — пыльцевход; 3 — покровы семязачатка; 4—семяножка; 5—макроспороцит; 6 —макроспоры; 7 — одноядерный зародышевый мешок; 8,9 — двухъядерные мешки; И,12 - молодой и зрелый восьмиядерные мешки; 13—яйцеклетка; 14 — синергиды; 15 — полярные ядра; 16 — антиподы.
Последний растет, его ядро трижды делится митотически, в результате чего образуется восемь дочерних ядер. Они располагаются по четыре двумя группами— вблизи, пыльцевхода зародышевого мешка и на противоположном полюсе. Затем от каждого полюса отходит но одному ядру в центр зародышевого мешка. Это так называемыеполярные ядра. В дальнейшем они могут сливаться, превращаясь в одно центральное, или вторичное диплоидное ядро (или их слияние происходит позднее, при оплодотворении). Остальные шесть ядер, по три на каждом полюсе, разделяются тонкими клеточными перегородками. При этом на полюсе у пыльцевхода образуется яйцевой аппарат, состоящий из яйцеклетки и двух клеток-синергид. На противоположном полюсе возникают так называемые клетки-антиподы, которые определенное время участвуют в доставке к клеткам зародышевого мешка питательных веществ, а затем исчезают'. Такая восьмиядерная семиклеточная структура — зародышевый мешок — является зрелым женским гаметофитом, готовым к оплодотворению. Образование пыльцы и зародышевого мешка у большинства растений завершается одновременно.
Опыление. У цветковых растений процессу оплодотворения предшествует опыление.
Опыление—это перенос пыльцы из пыльников тычинок на рыльце пестика. Различают два типа опыления: самоопыление и перекрестное опыление. Присамоопылении пыльцевые зерна переносятся на рыльце пестика того же цветка (ячмень, горох, тюльпан). У перекрестноопыляющихся растений осуществляется перенос пыльцы из тычинок цветков одною растения на рыльце пестика другого.
Наиболее часто перекрестное опыление осуществляется насекомыми и значительно реже ветром (береза, ольха, пырей, осоки), птицами, водой (водные растения).
В процессе длительной эволюции приспособление цветка к опылению насекомыми привело к формированию ярких, хорошо заметных, часто с приятным запахом цветков с нектарниками, вырабатывающими сладкую сахаристую жидкость. Кроме того, у таких растений образуется много пыльцы, которая служит кормом для ряда насекомых. Привлеченные яркой окраской или запахом цветка, насекомые, извлекая нектар из глубины цветка, касаются липкой или шероховатой поверхности пыльцевых зерен, которая прилипает к их телу. Перелетев на другой цветок, насекомое переносит часть пыльцы на рыльце пестика.
У цветков ветроопыляемых растений околоцветник отсутствует или плохо развит и не препятствует движению ветра; тычинки длинные, свисающие; пыльца сухая и мелкая, образуется в большом количестве; рыльца пестиков длинные, часто перистые. Большинство ветроопыляемых растений цветут до появления листьев, что облегчает опыление.
При перекрестном опылении, в отличие от самоопыления, у растений повышается уровень гетерозиготности потомства, что позволяет ему легче адаптироваться к постоянному изменению условий среды. В то же время самоопыление имеет одно существенное преимущество по сравнению с перекрестным: оно не зависит от погодных условий и посредников, поэтому осуществляется при любых условиях.
Оплодотворение. Попав на рыльце пестика, пыльцевое зерно начинает прорастать (рис. 8.20). Из Beгетативной клетки развивается длинная пыльцевая трубка, дорастающая по тканям столби ка до завязи и далее —до семязачатка. Из генеративной клетки к этому моменту образуются два спермия, которые спускаются в пыльцевую трубку. Рост пыльцевой трубки стимулируют ауксины, вырабатываемые пестиками, а к завязи она направляется в результате хемотропизма.
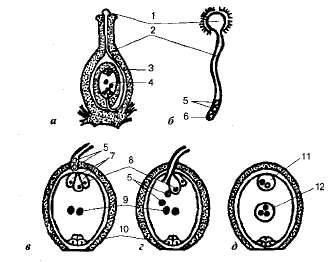
Рис. 8.20. Схема двойного оплодотворения у цветковых растений: а — продольный разрез пестика; б — прорастание пыльцевого зерна; в — проникновение пыльцевой трубки в зародышевый мешок; г — излияние содержимого пыльцевой трубки (двух спермиев) в зародышевый мешок; д — зародышевый мешок после оплодотворения: 1 — прорастающее пыльцевое зерно; 2 — пыльцевая трубка; 3 — завязь; 4 — зрелый зародышевый мешок; 5 — спермии; 6 — вегетативное ядро; 7 — си-нергиды; 8 — яйцеклетка; 9 — полярные ядра; 10 — антиподы; 11 — зигота; 12 — триплоидное ядро эндосперма.
Пыльцевая трубка входит в семязачаток через пыльцевход, ее ядро разрушается, а кончик трубки при соприкосновении с оболочкой зародышевого мешка разрывается, освобождая мужские гаметы. Спермии проникают в зародышевый мешок в синергиду или в щель между яйцеклеткой и центральным ядром. Вскоре после вхождения пыльцевой трубки в зародышевый мешок синергиды и антиподы отмирают.
После этого один из спермиев оплодотворяет яйцеклетку. В результате образуется диплоидная зигота, из которой развивается зародыш нового растительного организма. Второй спермий сливается с двумя полярными ядрами (или с центральным диплоидным ядром), образуя тришюидную клетку, из которой впоследствии возникает питательная ткань —эндосперм. В его клетках содержится запас питательных веществ, необходимых для развития зародыша растения.
Слияние одного спермия с яйцеклеткой, а другого с полярными ядрами представляет собой уникальную особенность покрытосеменных — двойное оплодотворение.Такой способ оплодотворения был открыт в 1898 г. русским цитологом и эмбриологом С. Г. Навашиным.
Благодаря двойному оплодотворению происходит очень быстрое образование и развитие эндосперма. В сочетании с огромным числом поколений этим достигается существенная экономия энергетических ресурсов растений. Двойное оплодотворение ускоряет также весь процесс формирования семязачатка и семени.
После оплодотворения семязачаток развивается в семя, завязь пестика формирует плод.
У многих растений в образовании плода участвуют и другие части цветка: разросшееся цветоложе, основания чашелистиков, лепестков, тычинок (например, у яблони, груши).