

МЕРИСТЕМЫ ИЛИ ОБРАЗОВАТЕ ЛЬНЫЕ ТКАНИ
Меристемы (от греч. «меристос» — делимый), или образовательные ткани, обладают способностью к делению и образованию новых клеток. За счет меристем формируются все прочие ткани и осуществляется длительный (в течение всей жизни) рост растения. У животных меристемы отсутствуют, чем объясняется ограниченный период их роста. Клетки меристем отличаются высокой метаболической активностью. Одни клетки меристем, получившие название инициальных, задерживаются на эмбриональной стадии развития в течение всей жизни растения, другие постепенно дифференцируются и превращаются в клетки различных постоянных тканей. Инициальная клетка меристемы принципиально может дать начало любой клетке организма. Тело наземных растений — производное относительно немногих инициальных клеток.
Первичные меристемы обладают меристематической активностью, т. е. способны к делению изначально. В ряде случаев способность к активному делению может вновь возникнуть и у клеток, уже почти утративших это свойство. Такие «вновь» возникшие меристемы называют вторичными.
В теле растения меристемы занимают различное положение, что позволяет их классифицировать. По положению в растении выделяют верхушечные, или апикальные (от лат. «апекс» — верхушка), боковые, или латеральные (от лат. «латус» — бок), и интеркалярные меристемы.
Апикальные меристемы располагаются на верхушках осевых органов растения и обеспечивают рост тела в длину, а латеральные — преимущественно рост в толщину. Каждый побег и корень, а также зародышевый корешок, почечка зародыша имеют апикальную меристему. Апикальные меристемы первичны и образуют конусы нарастания корня и побега (рис. 1).
Латеральные меристемы располагаются параллельно боковым поверхностям осевых органов, образуя своего рода цилиндры, на поперечных срезах имеющие вид колец. Часть из них относится к первичным. Первичными меристемами являются прокамбий и перицикл, вторичными — камбий и феллоген.
Интеркалярные, или вставочные, меристемы чаще первичны и сохраняются в виде отдельных участков в зонах активного роста (например, у оснований междоузлии, в основаниях черешков листьев).
Существуют также раневые меристемы. Они образуются в местах повреждения тканей и органов и дают начало каллюсу — особой ткани, состоящей из однородных паренхимных клеток, прикрывающие место поражения Каллюсо-образовательная способность растений используется в практике садоводства при размножении их черенками и прививками. Чем интенсивнее каллюсообразование, тем больше гарантия срастания подвоя с привоем и укоренения черенков. Образование каллюса— необходимое условие культуры тканей растения на искусственных средах.
Клетки апикальных меристем более или менее изодиаметричны по размерам и многогранны по форме. Межклетников между ними нет, оболочки тонкие, содержат мало целлюлозы. Полость клетки заполнена густой цитоплазмой с относительно крупным ядром, занимающим центральное положение. Вакуоли многочисленные, мелкие, но под световым микроскопом обычно не заметны. Эргастические вещества, как правило, отсутствуют. Пластид и митохондрий мало и они мелки.
Клетки боковых меристем различны по величине и форме. Они примерно соответствуют клеткам тех постоянных тканей, которые из них в дальнейшем возникают. Так, в камбии встречаются как паренхимные, так и прозенхимные инициали. Из паренхимных инициалей образуется паренхима проводящих тканей, а из прозенхимных — проводящие элементы.
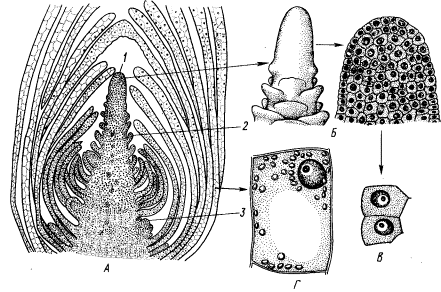
Рис. 1. Верхушечная меристема побега элодеи. А — продольный срез; 5 — конус нарастания (внешний вид и продольный срез); В — клетки первичной меристемы;
Г — паренхимная клетка сформировавшегося листа:
1 ~ конус нарастания, 2 — зачаток листа, 3 — бугорок пазушной почки
Онтогенетическая классификация меристем.
Все меристемы, функционирующие на протяжении всей жизни растения, можно подразделить на общие и специальные. К общим меристемам относится меристема зародыша, из которой развиваются апикальные (верхушечные) меристемы, а они, в свою очередь, дают начало третьей группе общих меристем — интеркалярным.
От апикальных меристем берут начало:
- протодерма, образующая эпидерму;
- основная меристема, дающая начало системе тканей основной паренхимы;
- прокамбий.
Клетки этих трёх тканей более вакуолизированы, чем апикальные меристемы, и обладают ограниченной способностью к делению, поэтому часто их называют полумеристемами.
Часть клеток прокамбия даёт начало первичным проводящим тканям — первичным ксилеме и флоэме; из других его клеток развивается камбий, из который, в свою очередь, образует вторичные проводящие ткани — вторичные ксилему и флоэму. Вторичные проводящие ткани образует и т. н. добавочный камбий, развивающийся не из прокамбия, а из постоянных тканей, живые клетки которых при этом дедифференцируются, то есть возвращаются из специализированного состояния к состоянию, близкому к эмбриональному, и вновь приобретают способность к делению.
Таким же образом в растении возникают и раневые меристемы, восстанавливающие повреждённые участки растения. На возможности новообразования меристем основано вегетативное размножение растений.
Кроме добавочного камбия, постоянные ткани также участвуют в образовании феллогена (пробкового камбия). Делясь периклинально (т.е. параллельно поверхности органа), клетки феллогена отделяют кнаружи будущие клетки пробки (феллемы), а внутрь — феллодерму