

С помощью хромосомных перестроек возможно:
- изучать взаимодействие генов при изменении их положения в хромосоме;
- выяснять влияние расположения эухроматинового и гетерохроматинового материала на фенотипический эффект гена;
- исследовать межхромосомные отношения в генотипе организма;
- получать новые группы сцепления.
Иначе говоря, ту структуру кариотипа и генотипа видов, которая отрабатывалась в ходе эволюции в течение сотен тысяч и миллионов лет, генетик имеет возможность перестроить в течение нескольких поколений. С помощью хромосомных перестроек можно создавать новые системы генотипов.
Хромосомные перестройки, происходящие как внутри одной хромосомы, так и между негомологичными хромосомами, являются очень важным механизмом перекомбинации генов внутри хромосомного набора каждого вида.
Из изложенного должно быть ясно, что перестройки хромосом могут изменять поведение хромосом в мейозе, действие генов, свойств доминирования генов, характер рекомбинации генов, гаметогенез и т. д. Поскольку естественный отбор контролирует все процессы в организме, очевидно, что потомство организмов с разными хромосомными перестройками будет иметь разные шансы на выживаемость.
Мы уже говорили о том, что явление гомологичной наследственной изменчивости объясняется в ряде случаев происхождением видов на основе хромосомных перестроек исходного кариотипа. Однако следует помнить, что когда формулировался закон гомологических рядов, то еще не было достаточного количества фактов о наличие хромосомных перестроек, и закон разрабатывался на основе фенотипической классификации наследственной изменчивости.
Теперь в генетике накоплено большое количество фактов, дающих основание признать, что одним из основных механизмов, обусловливающим возникновение гомологических рядов мутаций у близких видов, является процесс хромосомных перестроек. Транслокации, инверсии, дупликации и полиплоидия в процессе дифференциации вида на расы, подвиды и новые виды играют роль изолирующих факторов одной группы индивидуумов от другой. Указанные хромосомные перестройки вызывают нескрещиваемость особей в популяции, а также снижение плодовитости и жизнеспособности зигот вследствие нарушения генного баланса. Но в случаях возникновения жизнеспособной формы, гомозиготной по транслокации, инверсии или дупликации, она может оказаться приспособленной к определенным условиям существования и свободно размножится, а затем обособится в новый вид. У этого нового вида сохраняются прежние гены, но либо они окажутся в других группах сцепления, либо в иной последовательности расположения. Такие гены могут мутировать в том же направлении, что и у исходного вида, и таким образом обусловливать возникновение гомологичных рядов мутаций. Как показывают генетические исследования родственных видов, особенно в роде Drosophila, их генетические системы оказываются очень сходными, а различия касаются главным образом расположения отдельных генов.
Роль хромосомных перестроек важна и для эволюции генотипа. Как было показано, в результате транслокаций, дупликаций и инверсий гены вследствие эффекта положения изменяют характер Доминирования. Если полезная мутация гена является рецессивной, то с помощью эффекта положения она может проявиться в гетерозиготном состоянии и стабилизироваться в жизни вида. Значение транслокаций особенно велико в переносе отдельных участков аутосом на половые хромосомы. Эти перестройки являются важным фактором в определении нескрещиваемости видов животных.
Как мы уже знаем, Y-хромосома чаще состоит из гетерохроматина и генетически мало активна. Но у разных животных это проявляется в разной степени. Так, при общей малой активности Y-хромосомы дрозофилы один из ее участков, однако, является гомологичным участку в Х-хромосоме. В Y-хромосоме человека такой участок значительно длиннее, то же самое имеет место и у растения меландриум.
.jpg)
Сравнительное распределение генетически активных и инертных участков X- и Y- хромосом
В X- и Y-хромосомах могут быть как гомологичные участки, так и негомологичные, т. е. в Х-хромосоме всегда имеется свой, характерный для нее, район, отсутствующий в Y-хромосоме; гены в этом районе будут наследоваться сцепленно с полом. В Y-хромосоме также имеется район, не представленный в Х-хромосоме. Признаки, определяемые генами этого района (голандрическими генами), наследуются только по мужской линии при гетерогаметичности мужского пола.
Гомологичные и негомологичные участки в половых хромосомах, очевидно, могли произойти также посредством хромосомных перестроек. Об этом говорит тот факт, что у многих видов животных число гетероморфных хромосом сильно варьирует.
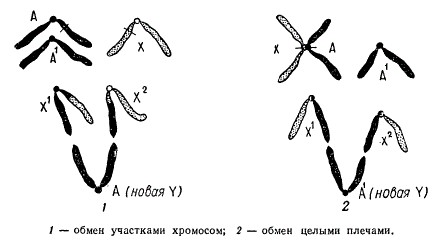
Схема образования гетероморфных хромосом у гетерогаметного пола (X0) в результате транслокации между X- хромосомой и аутосомой
Схематически показан ход возможного обмена фрагментами между Х-хромосомой и аутосомой у гетерогаметного пола Х0 в результате транслокации. Вследствие такой перестройки образуется новый тип гетерохромосом X1 X2 Y. Цитологический анализ мейоза у таких форм показывает образование тривалентов, что подтверждает правильность предположения о происхождении гетероморфных хромосом посредством транслокаций.
Цитологический анализ у некоторых родственных видов животных показывает, что гаплоидные числа хромосом в их наборе различаются, при этом, отдельные хромосомы могут нести гомологичные участки. У некоторых родственных видов дрозофилы известны 3 пары хромосом (D. willistoni), у других — 4 пары (D. melanogaster и D. americana), у третьей группы — 6 пар (D. virilis).
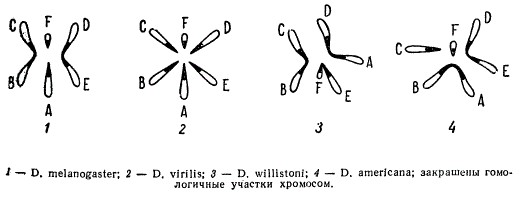
Гаплоидные наборы хромосом некоторых видов дрозофилы
Хромосомы могут превращаться в двуплечие хромосомы и, наоборот, может изменяться также и число хромосом в наборе.
Результаты исследования хромосомных перестроек убеждают во-первых, в наличии линейной дискретности хромосом и, Во-вторых, в том, что генотип представляет целостную систему, не сумму отдельных генов.
Рассмотрение хромосомных перестроек приводит к выводу, что они:
- лежат в основе изменений групп сцепления генов;
- изменяют характер наследования признаков и свойств в поколениях;
- изменяют проявление и взаимодействие генов;
- являются не только источником наследственной изменчивости комбинативного характера, но и механизмом преобразования генотипа и кариотипа в процессе эволюции;
- свидетельствуют о том, что многие гены, считавшиеся «классическими» точковыми мутациями, оказываются или дупликациями, или делециями, или инверсиями.
Хромосомные перестройки служат методом цитогенетической локализации генов в хромосомах, методом исследования механизма мейоза и тонкого картирования генов. Они могут быть использованы в практических целях для изменения групп сцепления генов, определяющих хозяйственно-ценные признаки.
Генетические и цитологические карты хромосом.
Генетически карты- граф-е изображение групп сцепления с указанием распол-я и расстояния м-у генами.В настоящее время карты групп сцепления построены для многих генетических объектов: от бактериофагов до человека. Осн-е этапы картир-я гена:
1. Опред. группы сцепления в кот. нах. картируемый ген.2. Опред. точной локализации гена в найденной группе сцепления.
Опре-е группы сцепления в к-й может нах-ся картир-й ген начинается с опр-я того, наход. он в аутосоме, либо сцеплен с полом. Это опр-е по рез-м рецепрокных скрещ-й. Если они совпадают, то ген расположен в аутосоме, если различны, то сцеплен с полом.
Генетические карты хромосом, схемы относительного расположения сцепленных между собой наследственных факторов — генов. Г. к. х. отображают реально существующий линейный порядок размещения генов в хромосомах и важны как в теоретических исследованиях, так и при проведении селекционной работы, т.к. позволяют сознательно подбирать пары признаков при скрещиваниях, а также предсказывать особенности наследования и проявления различных признаков у изучаемых организмов. Имея Г. к. х., можно по наследованию «сигнального» гена, тесно сцепленного с изучаемым, контролировать передачу потомству генов, обусловливающих развитие трудно анализируемых признаков; например, ген, определяющий эндосперм у кукурузы и находящийся в 9-й хромосоме, сцеплен с геном, определяющим пониженную жизнеспособность растения.
Гены, расположенные в одной хромосоме, в большинстве случаев наследуются совместно и образуют одну группу сцепления, количество которых, т. о., соответствует у каждого организма гаплоидному числу хромосом + кроссинговер.
Чем дальше друг от друга гены, тем выше вероятность кроссинговера и, следовательно, рекомбинации. Генетический анализ позволяет обнаружить перекрест только при различии гомологичных хромосом по составу генов, что при кроссинговере приводит к появлению новых генных комбинаций. Обычно расстояние между генами на Г. к. х. выражают как % кроссинговера (отношение числа мутантных особей, отличающихся от родителей иным сочетанием генов, к общему количеству изученных особей); единица этого расстояния — морганида — соответствует частоте кроссинговера в 1%.
Г. к. х. составляют для каждой пары гомологичных хромосом. Группы сцепления нумеруют последовательно, по мере их обнаружения. Кроме номера группы сцепления, указывают полные или сокращённые названия мутантных генов, их расстояния в морганидах от одного из концов хромосомы, принятого за нулевую точку, а также место центромеры. Составить Г. к. х. можно только для объектов, у которых изучено большое число мутантных генов. Например, у дрозофилы идентифицировано свыше 500 генов, локализованных в её 4 группах сцепления, у кукурузы — около 400 генов, распределенных в 10 группах сцепления. У менее изученных объектов число обнаруженных групп сцепления меньше гаплоидного числа хромосом. Так, у домовой мыши выявлено около 200 генов, образующих 15 групп сцепления (на самом деле их 20). У человека из ожидаемых 23 групп сцепления (23 пары хромосом) идентифицировано только 10, причём в каждой группе известно небольшое число генов; наиболее подробные карты составлены для половых хромосом. У бактерий, которые являются гаплоидными организмами, имеется одна, чаще всего непрерывная, кольцевая хромосома и все гены образуют одну группу сцепления. При переносе генетического материала из клетки-донора в клетку-реципиент, например при конъюгации, кольцевая хромосома разрывается и образующаяся линейная структура переносится из одной бактериальной клетки в другую (у кишечной палочки в течение 110—120 мин). Искусственно прерывая процесс конъюгации, можно по возникшим типам рекомбинантов установить, какие гены успели перейти в клетку-реципиент. В этом состоит один из методов построения Г. к. х. бактерий, детально разработанных у ряда видов. Ещё более детализированы Г. к. х. некоторых бактериофагов.
Цитологические карты хромосом, схематическое изображение хромосом с указанием мест фактического размещения отдельных генов, полученное с помощью цитологических методов. Ц. к. х. составляют для организмов, для которых обычно уже имеются генетические карты хромосом. Каждое место расположения гена (локус) на генетической карте организма, установленное на основе частоты перекреста участков хромосом (кроссинговера), на Ц. к. х. привязано к определённому, реально существующему участку хромосомы, что служит одним из основных доказательств хромосомной теории наследственности. Для построения Ц. к. х. используют данные анализа хромосомных перестроек (вставки, делеции и др.) и, сопоставляя изменения морфологических признаков хромосом при этих перестройках с изменениями генетических свойств организма, устанавливают место того или иного гена в хромосоме. Пользуясь методом хромосомных перестроек, амер. генетик К. Бриджес составил в 1935 подробную Ц. к. х. плодовой мушки дрозофилы, наиболее полно генетически изученного организма. Гигантские хромосомы насекомых отряда двукрылых оказались самыми удобными для построения Ц. к. х., т.к. наряду с большими размерами обладают чёткой морфологической очерченностью: каждый участок этих хромосом имеет свой определённый и чёткий рисунок, обусловленный характерным чередованием по длине ярко окрашиваемых участков (дисков) и слабо окрашиваемых (междисков). Цитологическими методами легко определить отсутствие участка хромосомы или перенос его в др. место. Сопоставление Ц. к. х. с генетическими показало, что физическое расстояние между генами в хромосомах не соответствует генетическому (видимо, частота кроссинговера неодинакова в разных участках хромосом), поэтому плотность распределения генов на цитологических и генетических картах хромосом различна. Так было установлено важное генетическое явление — неравномерность частот перекреста по длине хромосомы. Линейное расположение генов и их последовательность, установленные генетическими методами, подтверждаются Ц. к. х. Современные методы цитологии и генетики позволяют построить Ц. к. х. многих организмов, в том числе человека