
1. Понятие активного центра. Структура и свойства активных центров ферментов
При изучении механизма химической реакции, катализируемой ферментами, исследователя всегда интересует не только определение промежуточных и конечных продуктов и выяснение отдельных стадий реакции, но и природа тех функциональных групп в молекуле фермента, которые обеспечивают специфичность действия фермента на данный субстрат (субстраты) и высокую каталитическую активность. Речь идет, следовательно, о точном знании геометрии и третичной структуры фермента, а также химической природы того участка (участков) молекулыфермента, который обеспечивает высокую скорость каталитической реакции. Участвующие в ферментативных реакциях молекулы субстратов часто имеют небольшие размеры по сравнению с молекулами ферментов, поэтому было высказано предположение, что при образовании фермент-субстратных комплексов в непосредственный контакт с молекулой субстрата, очевидно, вступает ограниченная часть аминокислот пептидной цепи. Отсюда возникло представление об активном центре фермента. Под активным центром подразумевают уникальную комбинацию аминокислотных остатков в молекуле фермента, обеспечивающую непосредственное связывание ее с молекулой субстрата и прямое участие в акте катализа (рис. 4.2). Установлено, что у сложных ферментов в состав активного центра входят также просте-тические группы.
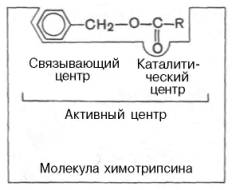
В активном центре условно различают так называемый каталитический центр, непосредственно вступающий в химическое взаимодействие с субстратом, и связывающий центр, или контактную («якорную») площадку, которая обеспечивает специфическое сродство к субстрату и формирование его комплекса с ферментом. В свою очередь молекула субстрата также содержит функционально различные участки: например, субстраты эстераз или протеиназ – одну специфическую связь (или группу атомов), подвергающуюся атаке со стороны фермента, и один или несколько участков, избирательно связываемых ферментом.
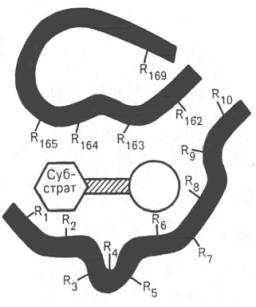
Рис. 4.2. Активный центр фермента (схема) (по Малеру и Кордесу).
Темные полосы - участки полипептидной цепи фермента; R - аминокислотные остатки и их порядковые номера (с N-конца).
Получены экспериментальные доказательства наличия в активном центре химотрипсина двух остатков гистидина и остатка серина, схематически представленных в трехмерной структурной модели предшественника этого фермента(рис. 4.3). Выявление химической природы и вероятной топографии групп активного центра – проблема первостепенной важности. Она сводится к определению природы аминокислот, их последовательности и взаиморасположения в активном центре. Для идентификации так называемых существенных аминокислотных остатков используют специфические ингибиторы ферментов (часто это субстратподобные вещества или аналоги коферментов), методы «мягкого» (ограниченного) гидролиза в сочетании с химической модификацией, включающей избирательное окисление, связывание, замещение остатков аминокислот и др.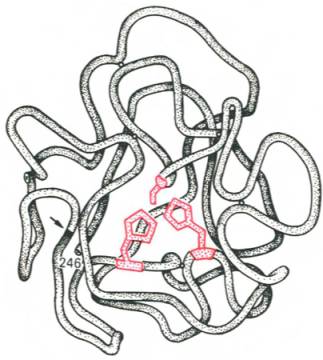
Рис. 4.3. Гипотетическая модель третичной структуры молекулы предшественника химотрипсина (по Ней-рату). Цветом выделены остатки серина и гисти-дина; стрелкой обозначено место отщепления N-концевого участка полипептидной цепи.
При помощи методов ингибиторного анализа были предприняты попытки установить закономерности состава и структуры активных центров у ферментов, относящихся к разным группам. В частности, при использованиидиизопропилфторфосфата (ДФФ), принадлежащего к так называемым нервным ядам, наблюдается полное выключение активного центра холинэстеразы – фермента, катализирующего гидролиз ацетилхолина на холин и уксусную кислоту. Оказалось, что этот ингибитор имеет близкое структурное сходство с ацетилхолином и подобно ему взаимодействует с ОН-группой остатка серина в активном центре. Вызывая фосфорилирование серина в активном центре ряда других ферментов, ДФФ также инактивирует их действие:
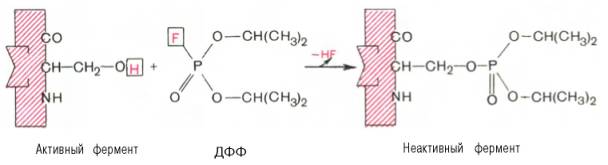
Показано, что ДФФ избирательно фосфорилирует в каждом чувствительном к нему ферменте только один остаток серина, наделенный функциональной активностью. Учитывая этот механизм действия ДФФ, сделаны попытки определения природы аминокислот в окружении «каталитического» остатка серина у ряда ферментов (табл. 4.2).
Из данных табл. 4.2 видно, что ферменты, сходные по типу действия, хотя и различаются специфичностью, могут иметь почти одинаковую последовательность аминокислотных остатков в тех участках, которые примыкают к остатку серина, несущему функционально активную гидро-ксильную группу. Существенное значение ОН-группысерина для акта катализа было доказано, кроме того, химическим ее блокированием или удалением, когда эстеразы полностью лишались ферментативной активности.

Предполагают, что формирование активного центра фермента начинается уже на ранних этапах синтеза белка-фермента (см. главу 14) на рибосоме, когда линейная одномерная структура пептидной цепи превращается в трехмерное тело строго определенной конфигурации. Образовавшийся белок приобретает информацию совершенно нового типа, а именно функциональную (в частности, каталитическую). Любые воздействия, приводящие к денатурации, т.е. нарушению третичной структуры, приводят к искажению или разрушению структуры активного центра и соответственно потере ферментом каталитических свойств. Если при подходящих внешних условиях удается восстановить нативную трехмерную структуру белка-фермента (ренатурировать его), то восстанавливается и его каталитическая активность. Это было показано впервые на примере рибо-нуклеазы поджелудочной железы (см. рис. 1.13).
Помимо активного центра, в молекуле фермента может присутствовать также аллостерический центр (или центры) (от греч. allos – другой, иной и steros – пространственный, структурный), представляющий собой участок молекулы фермента, с которым связываются определенные, обычно низкомолекулярные, вещества (эффекторы, или модификаторы), молекулы которых отличаются по структуре от субстратов. Присоединение эффектора к аллостерическому центру изменяет третичную и часто также четвертичную структуру молекулы фермента и соответственно конфигурацию активного центра, вызывая снижение или повышение энзимати-ческой активности. Ферменты, активность каталитического центра которых подвергается изменению под влиянием аллостерических эффекторов, связывающихся с аллостерическим центром, получили название аллосте-рических ферментов.
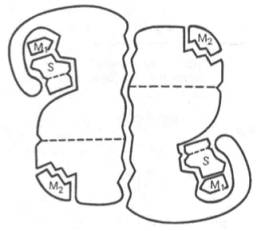
Рис. 4.4. Схематическое изображение аллостерического фермента, состоящего из двух протомеров, соединенных по типу гетерологи-ческой («голова»-«хвост») ассоциации (по Кошленду). S – субстрат; М1 – модификатор, связывающийся в активном центре; М2 – модификатор, связывающийся в аллосте-рическом центре (эффектор).
Отличительной особенностью ряда аллостерических ферментов является наличие в молекуле олигомерногофермента нескольких активных центров и нескольких аллостерических регуляторных центров, пространственно удаленных друг от друга. В аллостерическом ферменте каждый из двух симметрично построенных протомеров содержит один активный центр, связывающий субстрат S, и один аллостерический центр, связывающий эффектор М2, т.е. 2 центра в одной молекуле фермента (рис. 4.4). Получены доказательства, что для субстратааллостерические ферменты, помимо активного центра, содержат и так называемые эффекторные центры; при связывании с эффекторным центром субстрат не подвергается каталитическому превращению, однако он влияет на каталитическую эффективность активного центра. Подобные взаимодействия между центрами, связывающими лиганды одного типа, принято называть гомотропными взаимодействиями, а взаимодействия между центрами, связывающими лиганды разных типов, – гетеротропными взаимодействиями.
Таким образом, приведенные сведения о химической природе активного центра и аллостерических участках свидетельствуют о том, что в энзима-тическом катализе, как и в реакции связывания субстрата, участвует не ограниченная и небольшая часть фермента, как предполагалось ранее, а значительно большая часть молекулыбелка-фермента. Этими обстоятельствами, вероятнее всего, можно объяснить большие размеры и объемность трехмерной структуры молекулы фермента; эти же обстоятельства следует учитывать в программах создания искусственных низкомолекулярных аналогов ферментов (синзимов), обладающих свойствами на-тивных ферментов (см. ранее).
2. Структура и номенклатура нуклеотидов. Минорные нуклеозиды и нуклеотиды. Биологическая роль нуклеотидов
Строение[править | править вики-текст]
Нуклеотиды являются сложными эфирами нуклеозидов и фосфорных кислот. Нуклеозиды, в свою очередь, являются N-гликозидами, содержащими гетероциклический фрагмент, связанный через атом азота с C-1 атомом остатка сахара.
В природе наиболее распространены нуклеотиды, являющиеся β-N-гликозидами пуринов или пиримидинов и пентоз — D-рибозы или D-2-дезоксирибозы. В зависимости от структуры пентозы различают рибонуклеотиды и дезоксирибонуклеотиды, которые являются мономерами молекул сложных биологических полимеров (полинуклеотидов) — соответственно РНК или ДНК[1].
Фосфатный остаток в нуклеотидах обычно образует сложноэфирную связь с 2′-, 3′- или 5′-гидроксильными группами рибонуклеозидов, в случае 2′-дезоксинуклеозидов этерифицируются 3′- или 5′-гидроксильные группы.
Большинство нуклеотидов являются моноэфирами ортофосфорной кислоты, однако известны и диэфиры нуклеотидов, в которых этерифицированы два гидроксильных остатка — например, циклические нуклеотиды циклоаденин- и циклогуанин монофосфаты (цАМФ и цГМФ). Наряду с нуклеотидами — эфирами ортофосфорной кислоты (монофосфатами) в природе также распространены и моно- и диэфиры пирофосфорной кислоты (дифосфаты, например, аденозиндифосфат) и моноэфиры триполифосфорной кислоты (трифосфаты, например, аденозинтрифосфат).
Номенклатура[править | править вики-текст]
| Код | Означает | Комплементарная пара |
|---|---|---|
| A | A | T в ДНК; U в РНК |
| C | C | G |
| G | G | C |
| T или U |
T в ДНК; U в РНК |
A |
| M | A или C |
K |
| R | A или G |
Y |
| W | A или T |
W |
| S | C или G |
S |
| Y | C или T |
R |
| K | G или T |
M |
| V | A или C или G |
B |
| H | A или C или T |
D |
| D | A или G или T |
H |
| B | C или G или T |
V |
| X или N |
A или C или G или T (U) |
любой |
Соединения, состоящие из двух нуклеотидовых молекул, называются динуклеотидами, из трёх — тринуклеотидами, из небольшого числа — олигонуклеотидами, а из многих — полинуклеотидами, или нуклеиновыми кислотами.
Названия нуклеотидов представляют собой аббревиатуры в виде стандартных трёх- или четырёхбуквенных кодов.
Если аббревиатура начинается со строчной буквы «д» (англ. d), значит подразумевается дезоксирибонуклеотид; отсутствие буквы «д» означает рибонуклеотид. Если аббревиатура начинается со строчной буквы «ц» (англ. c), значит речь идёт о циклической форме нуклеотида (например, цАМФ).
Первая прописная буква аббревиатуры указывает на конкретное азотистое основание или группу возможных нуклеиновых оснований, вторая буква — на количество остатков фосфорной кислоты в структуре (М — моно-, Д — ди-, Т — три-), а третья прописная буква — всегда буква Ф («-фосфат»; англ. P).
Латинские и русские коды для нуклеиновых оснований:
- A — А: Аденин;
- G — Г: Гуанин;
- C — Ц: Цитозин;
- T — Т: Тимин (5-метилурацил), встречается у бактериофагов в ДНК, занимает место урацила в РНК;
- U — У: Урацил, встречается в РНК , занимает место тимина в ДНК.
Общепринятые буквенные коды для обозначения нуклеотидных оснований соответствуют номенклатуре, принятой Международным союзом теоретической и прикладной химии (International Union of Pure and Applied Chemistry, сокращённо — англ. IUPAC, ИЮПАК) и Международным союзом биохимии и молекулярной биологии (International Union of Biochemistry and Molecular Biology (англ.)русск., сокращённо — англ. IUBMB). Если при секвенировании последовательности ДНК или РНК возникает сомнение в точности определения того или иного нуклеотида, помимо пяти основных (A, C, T, G, U), используют другие буквы латинского алфавита в зависимости от того, какие наиболее вероятные нуклеотиды могут находиться в данной позиции последовательности. Эти же дополнительные буквы используют для обозначения вырожденных (не совпадающих у разных гомологичных последовательностей) позиций, например при записи последовательности праймеров для ПЦР.
Длину секвенированных участков ДНК (гена, сайта, хромосомы) или всего генома указывают в парах нуклеотидов (пн), или парах оснований (англ. base pairs, сокращённо bp), подразумевая под этим элементарную единицу двухцепочечной молекулы нуклеиновой кислоты, сложенную из двух спаренных комплементарных оснований.
Биологическая роль[править | править вики-текст]
1. Универсальный источник энергии (АТФ и его аналоги).
2. Являются активаторами и переносчиками мономеров в клетке(УДФ-глюкоза)
3. Выступают в роли коферментов (ФАД, ФМН, НАД+, НАДФ+)
4. Циклические мононуклеотиды являются вторичными посредниками при действии гормонов и других сигналов(цАМФ, цГМФ).
5. Аллостерические регуляторы активности ферментов.
6. Являются мономерами в составе нуклеиновых кислот, связанные 3′-5′-фосфодиэфирными связями.
3. Репликация ДНК: общая схема процесса репликации и его этапы
Реплика́ция (от лат. replicatio — возобновление) — процесс синтеза дочерней молекулы дезоксирибонуклеиновой кислоты на матрице родительской молекулы ДНК. В ходе последующего деления материнской клетки каждая дочерняя клетка получает по одной копии молекулы ДНК, которая является идентичной ДНК исходной материнской клетки. Этот процесс обеспечивает точную передачу генетической информации из поколения в поколение. Репликацию ДНК осуществляет сложный ферментный комплекс, состоящий из 15—20 различных белков, называемый реплисомой (англ.)[1].
Ферменты (хеликаза, топоизомераза) и ДНК-связывающие белки расплетают ДНК, удерживают матрицу в разведённом состоянии и вращают молекулу ДНК. Правильность репликации обеспечивается точным соответствием комплементарных пар оснований и активностью ДНК-полимеразы, способной распознать и исправить ошибку. Репликация у прокариот осуществляется несколькими разными ДНК-полимеразами. ДНК-полимераза I действует на запаздывающей цепи для удаления РНК-праймеров и дорепликации очищенных мест ДНК. ДНК полимераза III — основной фермент репликации ДНК, осуществляющий синтез ведущей цепи ДНК и фрагментов Оказаки при синтезе запаздывающей цепи. Далее происходит закручивание синтезированных молекул по принципу суперспирализации и дальнейшей компактизации ДНК. Синтез энергозатратный.
Цепи молекулы ДНК расходятся, образуют репликационную вилку, и каждая из них становится матрицей, на которой синтезируется новая комплементарная цепь. В результате образуются две новые двуспиральные молекулы ДНК, идентичные родительской молекуле.