

Коллаген
Характерным компонентом структуры соединительной ткани являются коллагеновые волокна. Они построены в основном из своеобразного белка - коллагена . Коллаген составляет 25-33% от общего количества белка организма взрослого человека или 6% от массы тела.
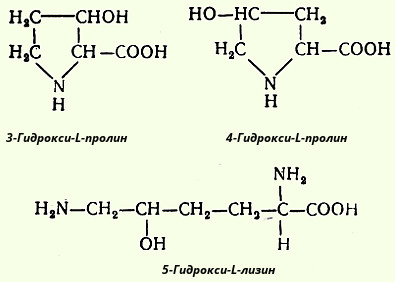
Видимые в оптическом микроскопе коллагеновые волокна состоят из различимых в электронном микроскопе фибрилл. Последние в свою очередь состоят из вытянутых в длину, соединенных между собой конец в конец белковых молекул, названных тропоколлагеном. Тропоколлаген содержит три полипептидные цепи, которые сливаются в спиралевидный триплет. Необходимо четко разграничить понятия "коллагеновые волокна" и "коллаген". Первое понятие является по-существу морфологическим и не может быть сведено к биохимическим представлениям о коллагене как белке. Коллагеновое волокно представляет собой гетерогенное образование и содержит, кроме белка коллагена, и другие химические компоненты. Молекула тропоколлагена - это белок коллаген. Одной из отличительных черт данного белка является то, что 1/3 всех его аминокислотных остатков составляет глицин, а 1/4-пролин, гидроксипролин и около 1 % - гидроксилизин. Некоторые молекулярные формы коллагена в своем составе имеют также 3-гидрокси-L-пролин, хотя и в весьма ограниченном количестве.
Каждая полипептидная цепь тропоколлагена содержит около 1000 аминокислотных остатков и имеет отн. мол. м. 120 000.
Дальнейшее изучение аминокислотного состава и последовательности чередования аминокислот в полипептидных цепях тропоколлагена показало, что существуют два типа цепей: цепи α1 и цепи α2. Кроме того, имеются четыре разновидности цепи αl: α1(I), αl(II), αl(III) и αl(IV). В табл. 61 представлены данные о структуре тропоколлагенов коллагеновых волокон различных тканей.
| Таблица 61. Тип коллагена и некоторые свойства (по Уайту и др., 1981) | |||
| Тип | Ткань | Полипептидные цепи | Дополнительная характеристика |
| I | Кожа, кость, связки | [α1 (I)]2 α2 | < 10 остатков гидрокcилизина в цепи |
| II | Хрящ | [α1 (II)]3 | > 10 остатков гидроксилизина в цепи |
| III | Кровеносные сосуды, кожа плода | [α1 (III)]3 | Слишком высокое содержание гидроксипролина и глицина |
| IV | Базальная мембрана | [α1 (IV)]3 | Высокое содержание 3-гидроксипролина; > 20 остатков гидроксилизина в цепи; низкое содержание аланина |
Как и все белки, коллаген синтезируется клетками из свободных аминокислот. Однако аминокислотные остатки, специфичные для молекулы коллагена, - гидроксипролил и гидроксилизид не образуются из соответствующих свободных аминокислот. Образование этих аминокислотных остатков происходит после включения пролина и лизина в полипептидную цепь с участием ферментов пролингидроксилазы или лизингидроксилазы и кофактора - аскорбиновой кислоты.
Эластин
Эластин основной белковый компонент, из которого состоят эластические волокна. Он отличается от коллагена по химическому составу и молекулярной структуре.
Общим для эластина и коллагена является большое содержание глицина и пролина, наличие гидроксипролина, хотя последнего в эластине примерно в 10 раз меньше, чем в коллагене. Как и в коллагене, в эластине нет цистеина, мало метионина и отсутствует триптофан.
В отличие от коллагена в эластине значительно больше валина и аланина и меньше глутаминовой кислоты и аргинина. В целом характерной особенностью первичной структуры эластина является слишком малое содержание полярных аминокислотных остатков. При ферментативном гидролизе эластина в гидролизате обнаруживаются десмозин и изодесмозин. Эти соединения содержатся только в эластине. Структура их довольно необычна: четыре остатка лизина, соединяясь своими R-группами, образуют замещенное пиридиновое кольцо.
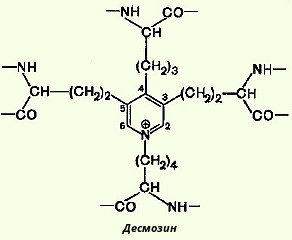
Есть основание считать, что при образовании десмозина сначала три остатка лизина окисляются до соответствующих ε-альдегидов, а затем происходит их соединение с четвертым остатком лизина.
Считают, что именно благодаря своей структуре десмозин и изодесмозин могут одновременно входить в состав четырех пептидных цепей. По-видимому, этим можно объяснить то, что эластин в отличие от других фибриллярных белков способен растягиваться в двух направлениях.
В гидролизатах эластина найдена еще одна необычная "аминокислота", пик которой на хроматограммах располагается между орнитином и лизином. Оказалось, что это - лизиннорлейцин, который обеспечивает наряду с десмозином и изодесмозином поперечные связи в молекуле эластина:
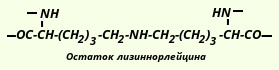
Эластин вместе с коллагеном, протеогликанами и рядом глико- и мукопротеинов является продуктом биосинтетической деятельности фибробластов. Непосредственным продуктом клеточного биосинтеза считается не эластин, а его предшественник - тропоэластин (в коллагене - проколлаген). Тропоэластин не содержит поперечных связей, он растворим. В последующем тропоэластин превращается в зрелый эластин, нерастворимый, содержащий большое количество поперечных связей. Десмозин, изодесмозин и лизиннорлейции, по-видимому, не исчерпывают список соединений, образующих поперечные связи в молекуле эластина.
Хондроитин-4-сульфат и хондроитин-6-сульфат построены по одному плану. Отличие между ними заключается в локализации сульфатной группы.
Несмотря на минимальные различия в химической структуре, физико-химические свойства хондроитин-4-сульфата и хондроитин-6-сульфата существенно отличные, а также различаются по своему преимущественному распределению среди разных видов соединительной ткани (табл. 63).
Дерматан-сульфат особенно характерен для дермы (кожи). Дерматан-сульфат резистентен к действию гиалуронидаз (тестикулярной и бактериальной). В этом одно из его отличий от хондроитин-сульфатов. Кроме того, в состав дисахаридной единицы дерматан-сульфата входит L-идуроновая, а не D-глюкуроновая кислота (в малом количестве D-глюкуроновую кислоту можно обнаружить в повторяющихся единицах дерматан-сульфата).
О биологической роли дерматан-сульфата почти ничего не известно. Роль этого гликозаминогликана не может быть сведена только к стабилизации коллагеновых пучков, так как дерматан-сульфат обнаруживается и в тканях эктодермального происхождения, не содержащих коллагена, например, в слизистой оболочке желудка.
Кератан-сульфат впервые был выделен из роговой оболочки глаза быка, отсюда и название этого гликозаминогликана. В противоположность всем остальным гликозаминогликанам кератан-сульфат не содержит ни D-глюкуроновой, ни L-идуроновой кислоты.
Установлено, что кератан-сульфат, выделенный из роговицы глаза (кератан-сульфат I), и кератан-сульфат, полученный из хрящевой ткани (кератан-сульфат II), отличаются по степени сульфатированности и строению связи между кератан-сульфатом и пептидной частью протеогликана.
Гепарин и гепарин-сульфат. Гепарин известен прежде всего как антикоагулянт. Однако его следует относить к гликозаминогликанам, так как он синтезируется тучными клетками, которые являются разновидностью клеточных элементов соединительной ткани. Он может входить в состав протеогликанов, с гликозаминогликанами его объединяет и химическая структура (см рис). Ряд исследователей считают, что точная структурная формула гепарина еше неизвестна.
Гликозаминогликаны - линейные отрицательно заряженные гетерополисахариды. РаНbше их называли мукополисахаридами, так как они обнаруживались в слизистых секретах (мукоза) и придавали этим секретам вязкие, смазочные свойства. Эти свойства обусловлены тем, что гликозаминогликаны могут связывать большие количества воды, в результате чего межклеточное вещество приобретает желеобразный характер.
Протеогликаны - высокомолекулярные соединения, состоящие из белка (5-10%) и гликозаминогликанов (90-95%). Они образуют основное вещество межклеточного матрикса соединительной ткани и могут составлять до 30% сухой массы ткани.
Белки в протеогликанах представлены одной полипептидной цепью разной молекулярной массы. Полисахаридные компоненты у разных протеогликанов разные. Протеогликаны отличаются от большой группы белков, которые называют гликопротеинами. Эти белки тоже содержат олигосахаридные цепи разной длины, ковалентно присоединённые к полипептидной основе. Углеводный компонент гликопротеинов гораздо меньше по массе, чем у протеогликанов, и составляет не более 40% от общей массы. Гликопротеины выполняют в организме человека разные функции и присутствуют во всех классах белков - ферментах, гормонах, транспортных, структурных белках и др. Представители гликопротеинов - коллаген и эластин, иммуноглобулины, ангиотензиноген, трансферрин, церулоплазмин, внутренний фактор Касла, тиреотропный гормон. Гликозаминогликаны и протеогликаны, являясь обязательными компонентами межклеточного матрикса, играют важную роль в межклеточных взаимодействиях, формировании и поддержании формы клеток и органов, образовании каркаса при формировании тканей. Благодаря особенностям своей структуры и физико-химическим свойствам, протеогликаны и гликозаминогликаны могут выполнять в организме человека следующие функции:
-
они являются структурными компонентами межклеточного матрикса;
-
протеогликаны и гликозаминогликаны специфически взаимодействуют с коллагеном, эластином, фибронектином, ламинином и другими белками межклеточного матрикса;
-
все протеогликаны и гликозаминогликаны, являясь полианионами, могут присоединять, кроме воды, большие количества катионов (Na+, K+, Са2+) и таким образом участвовать в формировании тургора различных тканей;
-
протеогликаны и гликозаминогликаны играют роль молекулярного сита в межклеточном матриксе, они препятствуют распространению патогенных микроорганизмов;
-
гиалуроновая кислота и протеогликаны выполняют рессорную функцию в суставных хрящах;
-
гепарансульфатсодержащие протеогликаны способствуют созданию фильтрационного барьера в почках;
-
кератансульфаты и дерматансульфаты обеспечивают прозрачность роговицы;
-
гепарин - антикоагулянт;
-
гепарансульфаты - компоненты плазматических мембран клеток, где они могут функционировать как рецепторы и участвовать в клеточной адгезии и межклеточных взаимодействиях. Они также выступают компонентами синаптических и других пузырьков.
Строение и классы гликозаминогликатов. Гликозаминогликаны представляют собой длинные неразветвлённые цепи гетерополиса-харидов. Они построены из повторяющихся дисахаридных единиц. Одним мономером этого дисахарида является гексуроновая кислота (D-глюкуроновая кислота или L-идуроновая), вторым мономером - производное аминосахара (глюкоз- или галактозамина). NH2-rpynna аминосахаров обычно ацетилирована, что приводит к исчезновению присущего им положительного заряда. Кроме гиалуроновой кислоты, все гликозаминогликаны содержат сульфатные группы в виде О-эфиров или N-сульфата.
Гиалуроновая кислота находится во многих органах и тканях. В хряще она связана с белком и участвует в образовании протеогликановых агрегатов, в некоторых органах (стекловидное тело глаза, пупочный канатик, суставная жидкость) встречается и в свободном виде. Предполагается, что в суставной жидкости гиалуроновая кислота выполняет роль смазочного вещества, уменьшая трение между суставными поверхностями. Повторяющаяся дисахаридная единица в гиалуроновой кислоте имеет следующую структуру:
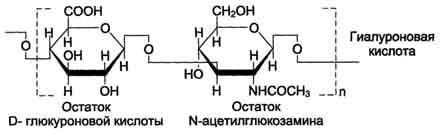
Гиалуроновая кислота содержит несколько тысяч дисахаридных единиц, молекулярная масса её достигает 105 - 107 Д.
Хондроитинсульфаты - самые распространённые гликозаминогликаны в организме человека; они содержатся в хряще, коже, сухожилиях, связках, артериях, роговице глаза. Хондроитинсульфаты являются важным составным компонентом агрекана - основного протеогликана хрящевого матрикса. В организме человека встречаются 2 вида хондроитинсульфатов: хондроитин-4-сульфат и хондроитин-6-сульфат. Они построены одинаковым образом, отличие касается только положения сульфатной группы в молекуле N-ацетилгалактозамина.
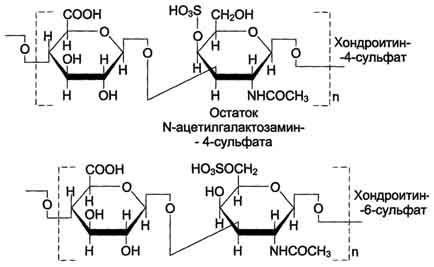
Одна полисахаридная цепь хондроитинсульфата содержит около 40 повторяющихся дисахаридных единиц и имеет молекулярную массу 104 - 106 Д.
Кератансульфаты - наиболее гетерогенные гликозаминогликаны; отличаются друг от друга по суммарному содержанию углеводов и распределению в разных тканях. Кератансульфат I находится в роговице глаза и содержит кроме повторяющейся дисахаридной единицы L-фукозу, D-маннозу и сиаловую кислоту. Кератансульфат II был обнаружен в хрящевой ткани, костях, межпозвоночных дисках. В его состав помимо Сахаров дисахаридной единицы входят N-ацетилгалактозамин, L-фукоза, D-манноза и сиаловая кислота. Кератансульфат II входит в состав агрекана и некоторых малых протеогликанов хрящевого матрикса. В отличие от других гликозаминогликанов, кератансульфаты вместо гексуроновой кислоты содержат остаток галактозы.
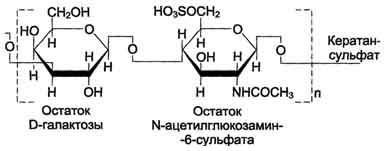
Молекулярная масса одной цепи кератансуль-фата колеблется от 4 × 103 до 20 × 103 Д.
Дерматансульфат широко распространён в тканях животных, особенно он характерен для кожи, кровеносных сосудов, сердечных клапанов. В составе малых протеогликанов (бигликана и декорина) дерматансульфат содержится в межклеточном веществе хрящей, межпозвоночных дисков, менисков. Повторяющаяся дисахаридная единица дерматансульфата имеет следующую структуру .
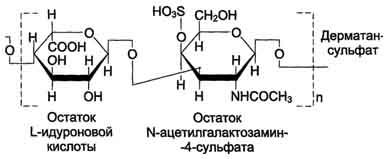
Молекулярная масса одной цепи дерматансульфата колеблется от 15 × 103 до 40 × 103 Д.
Гепарин - важный компонент противосвёртывающей системы крови (его применяют как антикоагулянт при лечении тромбозов). Он синтезируется тучными клетками и находится в гранулах внутри этих клеток. Наибольшие количества гепарина обнаруживаются в лёгких, печени и коже. Дисахаридная единица гепарина похожа на дисахаридную единицу гепарансульфата. Отличие этих гликозаминогликанов заключается в том, что в гепарине больше N-сульфатных групп, а в гепарансульфате больше N-ацетильных групп. Молекулярная масса гепарина колеблется от 6 × 103 до 25 × 103 Д
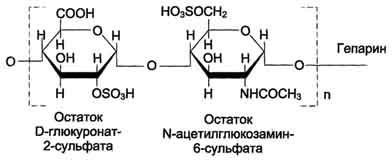
Гепарансульфат находится во многих органах и тканях. Он входит в состав протеогликанов базальных мембран. Гепарансульфат является постоянным компонентом клеточной поверхности. Структура дисахаридной единицы гепарансульфата такая же, как у гепарина. Молекулярная масса цепи гепарансульфата колеблется от 5 × 103 до 12 × 103 Д.
Коллагены - семейство родственных фибриллярных белков, секретируемых клетками соединительной ткани. Коллагены - самые распространённые белки не только межклеточного матрикса, но и организма в целом, они составляют около 1/4 всех белков организма человека. В межклеточном матриксе молекулы коллагена образуют полимеры, называемые фибриллами коллагена (более подробно это описано в разделе 15). Фибриллы коллагена обладают огромной прочностью и практически нерастяжимы. Они могут выдерживать нагрузку, в 10 000 раз превышающую их собственный вес. По прочности коллагеновые фибриллы превосходят прочность стальной проволоки того же сечения. Именно поэтому большое количество коллагеновых волокон, состоящих из коллагеновых фибрилл, входит в состав кожи, сухожилий, хрящей и костей.
Необычные механические свойства коллагенов связаны с их первичной и пространственной структурами. Молекулы коллагена состоят из трёх полипептидных цепей, называемых α-цепями. Идентифицировано более 20 α-цепей, большинство которых имеет в своём составе 1000 аминокислотных остатков, но цепи несколько отличаются аминокислотной последовательностью. В состав коллагенов могут входить три одинаковые или разные цепи.
Первичная структура α-цепей коллагена необычна, так капи, перевиваясь друг около друга, образуют трёхце-почечную правозакрученную суперспиральную молекулу, часто называемую тропоколлагеном. Цепи удерживаются друг около друга за счёт водородных связей, возникающих между амино- и карбоксильными группами пептидного остова разных полипептидных цепей, входящих в состав трёхспиральной молекулы. "Жёсткие" аминокислоты - пролин и гидроксипролин - ограничивают вращение полипептидного стержня и увеличивают тем самым стабильность тройной спирали. Глицин, имеющий вместо радикала атом водорода, всегда находится в месте пересечения цепей; отсутствие радикала позволяет цепям плотно прилегать друг к другу.
В результате такого скручивания пептидных остовов полипептидных цепей и наличия удлинённой структуры два других радикала из триады аминокислот Гли-X-Y оказываются на наружной поверхности молекулы тропоколлагена. Некоторые комплементарные участки молекул тропоколлагена могут объединяться друг с другом, формируя коллагеновые фибриллы, причём эти участки расположены таким образом, что одна нить тропоколлагена сдвинута по отношению к другой примерно на 1/4. Между радикалами аминокислот возникают ионные, водородные и гидрофобные связи.
Важную роль в формировании коллагеновых фибрилл играют модифицированные аминокислоты: гидроксипролин и гидроксилизин. Гидроксильные группы гидроксипролина соседних цепей тропоколлагена образуют водородные связи, укрепляющие структуру коллагеновых фибрилл. Радикалы лизина и гидроксилизина необходимы для образования прочных поперечных сшивок между молекулами тропоколлагена, ещё сильнее укрепляющие структуру коллагеновых фибрилл. Кроме того, к гидроксильной группе гидроксилизина могут присоединяться углеводные остатки (гликозилирование коллагена), функция которых пока неясна.
Таким образом, аминокислотная последовательность полипептидных цепей коллагена позволяет сформировать уникальную по своим механическим свойствам структуру, обладающую огромной прочностью. Изменение в первичной структуре коллагена может приводить к развитию наследственных болезней
Синтез и созревание коллагена - сложный многоэтапный процесс, начинающийся в клетке, а завершающийся в межклеточном матриксе. Синтез и созревание коллагена включают в себя целый ряд посттрансляционных изменений :
-
гидроксилирование пролина и лизина с образованием гидроксипролина (Hyp) и гидроксилизина (Hyl);
-
гликозилирование гидроксилизина;
-
частичный протеолиз - отщепление "сигнального" пептида, а также N- и С-концевых пропептидов;
-
образование тройной спирали.
Синтез полипептидных цепей коллагена
Полипептидные цепи коллагена синтезируются на полирибосомах, связанных с мембранами ЭР, в виде более длинных, чем зрелые цепи, предшественников - препро-α-цепей. У этих предшественников имеется гидрофобный "сигнальный" пептид на N-конце, содержащий около 100 аминокислот.
Основная функция сигнального пептида - ориентация синтеза пептидных цепей в полость ЭР. После выполнения этой функции сигнальный пептид сразу же отщепляется. Синтезированная молекула проколлагена содержит дополнительные участки - N- и С-концевые пропептиды, имеющие около 100 и 250 аминокислот, соответственно. В состав пропептидов входят остатки цистеина, которые образуют внутри- и межцепочечные (только в С-пептидах) S-S-связи. Концевые пропептиды не образуют тройную спираль, а формируют глобулярные домены. Отсутствие N- и С- концевых пептидов в структуре проколлагена нарушает правильное формирование тройной спирали.
ОСНОВНЫЕ СТРУКТУРЫ, ОБРАЗУЕМЫЕ КОЛЛАГЕНОМ
| Структура | Тип коллагена |
| Фибриллы Ассоциированные с фибриллами Сети Микрофибриллы «Заякоренные» фибриллы Трансмембранные домены Другие | I, II, III, V, XI IX, XII, XIV, XVI, XIX IV, VIII, X VI VII XIII, XVII XV, XVIII |
Фибриллообразующие (I, II, III, V и XI) типы
95% всего коллагена в организме человека составляют коллагены I, II и III типов, которые образуют очень прочные фибриллы. Они являются основными структурными компонентами органов и тканей, которые испытывают постоянную или периодическую механическую нагрузку (кости, сухожилия, хрящи, межпозвоночные диски, кровеносные сосуды), а также участвуют в образовании стромы паренхиматозных органов. Поэтому коллагены I, II и III типов часто называют интерстициальными. Во всех минерализующихся мезенхимных тканях присутствует коллаген I типа. Он беден гидроксилизином, слабо гликозилируется, образу широкие фибриллы. Коллаген II типа, наоборот, богат гидроксилизином сильно гидроксилирован. Коллаген III типа содержит большое количество остатков гидроксипролина и имеет межцепочечные дисульфидные связи. В отличие от коллагена I типа он не способен минерализоваться.
К классу фибриллообразующих относят также минорные коллагены V и XI типов. Основа структурной организации коллагеновых фибрилл - ступенчато расположенные параллельные ряды молекул тропоколлaгeна, которые сдвинуты на 1/4 относительно друг друга. Молекулы коллагена не связаны между собой «конец в конец», а между ними имеется промежуток в 35-40 нм. Предполагается, что в костной ткани эти промежутки выполняют роль центров минерализации, где откладываются кристаллы фосфата кальция. При электронной микроскопии фиксированные и контрастированные фибриллы коллагена выглядят поперечно исчерченными с периодом 67 нм, который включает одну тёмную и одну светлую полоски. Считают, что такое строение максимально повышает сопротивление всего агрегата растягивающим нагрузкам.
Фибриллы коллагена образуются спонтанно, путём самосборки. Но эти фибриллы ещё не являются зрелыми, так как не обладают достаточной прочностью (известно, что зрелое коллагеновое волокно толщиной в 1 мм выдерживает нагрузку до 10 кг).
Образовавшиеся коллагеновые фибриллы укрепляются внутри- и межцепочечными ковалентными сшивками (они встречаются только в коллагене и эластине). Эти сшивки образуются следующим образом:
- внеклеточный медьсодержащий фермент лизилоксидаза осуществляет окислительное дезаминирование е-аминогрупп в некоторых остатках лизина и гидроксилизина с образованием реактивных альдегидов (аллизина и гидроксиаллизина). Для этих реакций необходимо присутствие витаминов РР и В6.
- образовавшиеся реактивные альдегиды участвуют в формировании ковалентных связей между собой, а также с другими остатками лизина или гидроксилизина соседних молекул тропоколлагена, и в результате возникают поперечные «Лиз-Лиз-сшивки», стабилизирующие фибриллы коллагена.
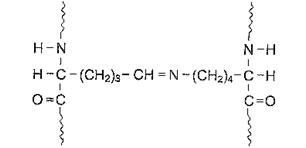 |
| Рис. 1.2. Шиффовы основания, образованные из боковых цепей лизина и аллизина. |
Шиффовы основания более часто встречаются в сухожилиях, а альдольная конденсация характерна для костей и зубов. Около 25% молекул тропоколлагена распадается, не образуя фибрилл. Получившиеся фрагменты выполняют сигнальные функции и стимулируют коллагеногенез. Количество поперечных связей в фибриллах коллагена зависит от функции и возраста ткани. Например, между молекулами коллагена ахиллова сухожилия сшивок особенно много, так как для этой структуры важна большая прочность. С возрастом количеств поперечных связей в фибриллах коллагена возрастает, что приводит к замедлению скорости его обмена у пожилых и старых людей.
При снижении активности лизилоксидазы, а также при недостатке меди или витаминов РР или В6 нарушается образование поперечных сшивок и, как следствие, снижаются прочность и упругость коллагеновых волокон. Такие структуры, как кожа, сухожилия, кровеносные сосуды, становятся хрупкими, легко разрываются.
Коллагены, ассоциированные с фибриллами
Этот класс объединяет коллагены, которые выполняют очень важную функцию: они ограничивают размер фибрилл, образуемых интерстициальными коллагенами (прежде всего, 1 и II типов), и участвуют в организации межклеточного матрикса в костях, коже, хрящах, сухожилиях. К этим коллагенам относят коллагены IX, XII, XIV и XVI типов. Коллагены этого класса сами фибрилл не формируют, но непосредственно связаны с фибриллами, которые образуют интерстициальные коллагены. К особенностям этого типа коллагенов относят наличие большого количества положительно заряженных групп, к которым могут присоединяться отрицательно заряженные гликозаминогликаны, например, гиалуроновая кислота и хондроитин-сульфат. Это обеспечивает их участие в организации межклеточного матрикса в хряще.
Коллагены, образующие сетеподобные структуры
К. этому классу относят коллагены IV, VIII, X типов.
Особенностью коллагена IV типа, структурного компонента базальных мембран, является то, что повторяющиеся спирализованные участки с последовательностью (Гли-х-у) часто прерываются короткими неспиральными сегментами. Это, вероятно, увеличивает гибкость коллагена IV типа и способствует образованию на его основе сетчатых структур.
Молекулы этого коллагена не могут ассоциироваться латерально с образованием фибрилл, так как N- и С-концевые пропептиды у него не отщепляются. Но именно эти фрагменты участвуют в образовании олигомерных форм коллагена, так как они имеют ряд потенциальных мест связывания (остатки цистеина и лизина). Дисульфидные мостики и поперечные лизиновые связи стабилизируют образующиеся олигомеры. Кроме этого, возможны латеральные взаимодействия спирализованных участков разных молекул с образованием суперспиралей. В базальной мембране из этих компонентов формируется сетчатая структура с гексагональными ячейками размером 170 нм.
Коллагены, образующие микрофибриллы
К этому классу относят коллаген VI типа, который является короткоцепочечным белком. Он образует микрофибриллы, которые располагаются между крупными фибриллами интерстициальных коллагенов. Этот коллаген широко представлен в хрящевом матриксе, но больше всего его содержится в межпозвоночных дисках. Две молекулы этого коллагена соединяются антипараллельно с образованием димера. Из димеров образуются тетрамеры, которые секретируются из клетки, и вне клетки связываются «конец в конец» с образованием микрофибрилл
Функции коллагена VI типа пока полностью неясны, хотя известно, что его микрофибриллы могут связываться со многими компонентами межклеточного матрикса: фибриллами интерстициальных коллагенов, гиалуроновой кислотой, протеогликанами. Молекула этого коллагена содержит многочисленные последовательности Apr-Гли-Асп (RGD), поэтому возможно его участие в клеточной адгезии через присоединение к мембранным адгезивным молекулам, например интегринам.
Коллагены, образующие «заякоренные» фибриллы
К этому классу относят коллагены VII и XVII типов, которые называют также коллагенами, связанными с эпителием, так как они обычно находятся в местах соединения эпителия с субэпителиальными слоями.
Коллаген VII типа - основной структурный компонент «заякоренных» фибрилл. Эти фибриллы играют важную роль в присоединении эпидермиса к дерме, так как одним концом они могут присоединяться к lamina densa, на которой лежит кожный эпителий, а другой их конец проникает в более глубокие субэпидермальные слои кожи и связывается там со структурами, называемыми «якорные диски».