

ВЗАИМОДЕЙСТВИЕ ГЕНОВ
Большое количество признаков формируется при участии нескольких генов, взаимодействие которых существенным образом отражается на особенностях фенотипа и приводит к отклонению от менделевской закономерности расщепления фенотипов. Описано несколько типов взаимодействия неаллельных генов: комплементарность, эпистази полимерия.
Комплементарность — взаимодействие неаллельных генов, которые обусловливают развитие нового признака, отсутствующего у родителей. Примером комплементарного действия у человека могут служить случаи, когда у глухих родителей рождаются дети с нормальным слухом.
Развитие нормального слуха находится под генетическим контролем десятков различных неаллельных генов, гомозиготное рецессивное состояние одного из которых может приводить к одной из форм наследственной глухоты. Таких форм у человека известно более 30. Если один из родителей является гомозиготой по рецессивному гену аа , а другой — гомозиготой по другому рецессивному гену bb, то все их дети будут двойными гетерозиготами и, следовательно, слышащими, поскольку доминантные аллели будут взаимно дополнять друг друга. Таким образом формируется новый по отношению к родителям признак — нормальный слух.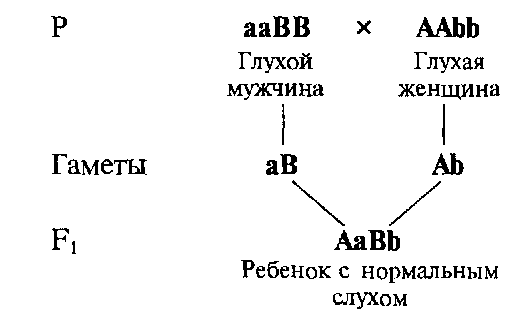
Схема, поясняющая возможность рождения детей с нормальным слухом у глухих родителей с различными генетическими формами глухоты
Эпистаз — взаимодействие неаллельных генов, при котором наблюдается подавление проявления одного гена действием другого, неаллельного гена. Подавляющий ген называется геном-супрессором, а подавляемый — гипостатическим геном. По-видимому, действие гена-супрессора на подавляемый ген сходно с принципом доминантность — рецессивность. Но существенное различие заключается в том, что эти гены не являются аллельными, т.е. расположены в негомологичных хромосомах или занимают различные локусы в гомологичных. Различают доминантный и рецессивный эпистаз. При доминантном эпистазе доминантный аллель гена-супрессора подавляет проявление доминантного аллеля другого гипостатического гена. При рецессивном эпистазе, или криптомерии, рецессивный аллель гена-супрессора, будучи в гомозиготном состоянии, не дает проявиться доминантной или рецессивным аллелям других генов.
Примером рецессивного эпистаза у человека может служить так называемый бомбейский фенотип, когда индивид, имеющий доминантный аллель группы крови системы АВ0 (например, аллель В, определяющий принадлежность человека к III или IV группе), идентифицируется в реакции агглютинации как человек с 0(I). Это состояние возникает в результате того, что данный индивид является рецессивной гомозиготой (hh) по другой, нежели система АВ0, генетической системе Hh. Для реализации аллелей IА и IЕ необходимо присутствие хотя бы одного доминантного I аллеля Н.
Полимерия — вид взаимодействия, когда эффекты нескольких неаллельных генов, определяющих один и тот же признак, примерно одинаковы. Подобные признаки получили название количественных, или полимерных признаков. Как правило, степень проявления полимерных признаков зависит от числа доминантных генов. Наследование полимерных признаков было впервые описано шведским генетиком Г. Нильсон-Эле в 1908 г. Проводя скрещивание различных форм пшеницы (с красными и белыми зернами), он наблюдал расщепление в F2 признака окраски в отношении: 15/16 (окрашенных и '/16 белых. Среди окрашенных зерен он наблюдал все переходы — от интенсивно окрашенных до слабо окрашенных.
Анализ особенностей расщепления показал, что в данном случае окраску зерен определяют два доминантных аллеля двух различных генов, а сочетания их рецессивных аллелей определяют отсутствие окраски. Поскольку полимерные гены имеют однонаправленное действие, их, как правило, обозначают одинаковыми буквами. Таким образом, исходные родительские формы имели генотипы А]А1А2А2 и а1а1а2а2. Наличие всех четырех доминантных аллелей определяло самую интенсивную окраску, трех доминантных аллелей (типа А1А1А2А2) — менее интенсивную окраску и т.д.
Примером полимерного наследования у человека является наследование окраски кожных покровов. В браке индивида негроидной расы (коренного жителя Африки) с черной окраской кожи и представителем европеоидной расы с белой кожей дети рождаются с промежуточным цветом кожи (мулаты). В браке двух мулатов потомки могут обладать любой окраской кожи: от черной до белой, поскольку пигментация кожи обусловлена действием трех или четырех неаллельных генов. Влияние каждого из этих генов на окраску кожи примерно одинаково.
Полимерное наследование характерно для так называемых количественных признаков, таких, как рост, вес, окраска кожных покровов, скорость протекания биохимических реакций, артериальное давление, содержание сахара в крови, особенности нервной системы, уровень интеллекта, и многих других, которые нельзя разложить на четкие фенотипические классы. Чем большее число неаллельных генов контролируют развитие количественного признака, тем менее заметны переходы между фенотипическими классами.