
Вопрос 1 Белки. Классификация белков. Уровни структурной организации белка. Химические связи, ответственные за формирование структуры белка. Биомедицинское значение полипептидов.
УРОВНИ СТРУКТУРНОЙ ОРГАНИЗАЦИИ БЕЛКА
Первичная структура
Под первичной структурой, уже знакомой нам из главы о пептидах (гл. 4), понимается последовательность аминокислот в полипептидной цепи (или цепях) и положение дисульфидных связей, если они имеются.
Вторичная структура
На этом структурном уровне описываются стерические взаимосвязи между расположенными близко друг к другу вдоль цепи аминокислотами. Вторичная структура может быть регулярной (а-спираль, складчатый  -слой) или не обнаруживать никаких признаков регулярности (неупорядоченная конформация).
-слой) или не обнаруживать никаких признаков регулярности (неупорядоченная конформация).
Третичная структура
Общее расположение, взаимную укладку различных областей, доменов и отдельных аминокислотных остатков одиночной полипептидной цепи называют третичной структурой данного белка. Четкой границы между вторичной и третичной структурами провести нельзя, однако под третичной структурой понимают стерические взаимосвязи между аминокислотными остатками, далеко отстоящими друг от друга по цепи.
Четвертичная структура
Если белки состоят из двух и более полипептидных цепей, связанных между собой нековалентными (не пептидными и не дисульфидными) связями, то говорят, что они обладают четвертичной структурой. Такие агрегаты стабилизируются водородными связями и электростатическими взаимодействиями между остатками, находящимися на поверхности полипептидных цепей. Подобные белки называют олигомерами, а составляющие их индивидуальные полипептидные цепи—протомерами, мономерами или субъединицами.
Многие олигомерные белки содержат два или четыре протомера и называются димерами или тетрамерами соответственно. Довольно часто встречаются олигомеры, содержащие более четырех протомеров, особенно среди регуляторных белков (пример — транскарбамоилаза). Олигомерные белки играют особую роль во внутриклеточной регуляции: их протомеры могут слегка менять взаимную ориентацию, что приводит к изменению свойств олигомера. Наиболее изученный пример — гемоглобин (гл. 16).
Роль первичной структуры в формировании более высоких уровней структурной организации белка
Вторичная и третичная структуры белка формируются самопроизвольно и определяются первичной структурой его полипептидной цепи. Параллельно синтезу цепи происходят ее локальное свертывание (образование вторичной структуры) и специфическая агрегация свернутых участков (формирование третичной структуры). Эти процессы детерминируются химическими группами, отходящими от атомов а-углерода соответствующих остатков. Например, обработка мономерного фермента рибонуклеазы мягким восстанавливающим агентом ф-меркаптоэтанолом) и денатурирующим агентом (мочевиной или гуанидином; см. ниже) приводит к инактивации белка и переходу его в неупорядоченную конформацию. Если медленно удалять денатурирующий агент и осуществлять постепенное реокисление, то вновь образуются S-S-связи и практически восстанавливается ферментативная активность. Нет никаких оснований думать, что существует независимый генетический контроль за формированием уровней структурной организации белка выше первичного, поскольку первичная структура специфически определяет и вторичную, и третичную, и четвертичную структуру (если она имеется)—т.е. конформацию белка. Нативной конформацией белка, в частности рибонуклеазы, по-видимому, является термодинамически наиболее устойчивая структура в данных условиях, т.е. при данных гидрофильных и гидрофобных свойствах среды.
Структура белка после его синтеза может модифицироваться (посттрансляционный процессинг); так, часто наблюдается превращение препрофермента в каталически активную форму или удаление «лидерной» последовательности, детерминирующей транспорт белков через мембраны (гл. 42).
Макромолекулярные белковые комплексы
Полифункциональные макромолекулярные комплексы, образующиеся в результате агрегации различных функциональных белков, каждый из которых обладает всеми четырьмя уровнями структурной организации, функционируют в цепи транспорта электронов (гл. 12), участвуют в биосинтезе жирных кислот (гл. 23) и метаболизме пирувата (гл. 18).
БИОМЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Пептиды имеют очень большое биомедицинское значение; особенно велика их роль в эндокринологии. Пептидами являются многие важнейшие гормоны человека. Их часто назначают больным для коррекции соответствующей недостаточности. Самый известный пример — введение инсулина больным сахарным диабетом. Пептидами являются также различные антибиотики (валиномицин, грамицидин А) и некоторые противоопухолевые препараты (например, блеомицин). Разработанные в последние годы методы быстрогохимического синтеза пептидов позволили наладить производство пептидных гормонов в значительных количествах; это разрешило многие проблемы, поскольку обычно гормоны присутствуют в организме животных в очень малых концентрациях и их трудно выделить в количествах, достаточных для терапевтических целей. По той же технологии осуществляется синтез и других пептидов, которые ввиду их малого содержания тоже трудно выделять из природных источников; в частности, это относится к вирусным пептидам, используемым в качестве вакцин.
Гемсодержащие белки участвуют в процессах связывания и транспорта кислорода, в транспорте электронов в фотосинтезе. Детальное изучение гемоглобина выявляет ряд структурных аспектов, общих для многих белков. Говоря о большом биомедицинском значении этих белков, мы имеем в виду, что результаты, полученные при исследовании, наглядно иллюстрируют структурно-функциональные взаимосвязи. Кроме того, эти исследования выявляют молекулярную основу ряда генетических болезней, таких как серповидноклеточная анемия (возникающая в результате изменения свойств поверхности b-субъединицы гемоглобина) или талассемия (хроническое наследуемое гемолитическое заболевание, характеризующееся нарушениями процессов синтеза гемоглобина). Летальный эффект цианида и окиси углерода объясняется тем, что эти вещества блокируют физиологическую функцию гемопротеинов - цитохромоксидазы и гемоглобина соответственно. Наконец, стабилизация четвертичной структуры дезоксигемоглобина 2,3-бифосфоглицератом (ДФГ) занимает центральное место в исследовании механизмов кислородной недостаточности в условиях высокогорья и процессов адаптации к этим условиям. [1]
ХИМИЧЕСКОЕ СТРОЕНИЕ
Химически гемоглобин относится к группе хромопротеидов. Его простетическая группа представляет собой ферросоединение протопорфирина IХ, с молекулярным составом С34Н32О4N4Fe и носит название гем (рис.1). Она придает соединению окраску. Белковый компонент гемоглобина называется глобином. Гемоглобиновая молекула содержит 4 гема и 1 глобин. Аминокислоты расположены в глобине в виде четырех полипептидных цепочек; две из них идентичны по структуре, и их обозначают как альфа-цепочки; две другие тоже идентичны между собой и их обозначают как бета-цепочки. Следовательно, формулу глобина можно выразить как альфа-альфа/бета-бета или альфа2бета2. альфа-полипептидная цепь состоит из 141, бета-полипептидная цепь - из 146 аминокислот.
Аминокислотный состав и последовательность (секвенция) альфа- и бета-цепей показаны на рис. 93. альфа-полипептидная цепь заканчивается комбинацией аминокислот валина-лейцина, а бета-полипептидная цепь - комбинацией валина-гистидина-лейцина. альфа- и бета-полипептидные цепи в гемоглобиновой молекуле не расположены линейно, как это выглядит на первый взгляд из данных ("первичная структура") на рис. 2. По причине существования интрамолекулярных сил, полипептидные цепи скручиваются в форме типичной для белков альфа-геликсовой спирали ("вторичная структура"). Сама альфа-геликсовая спираль на каждую альфа- и бета-полипептидную цепь огибается
пространственно, образуя сплетения овоидной формы ("третичная структура"). На рис. 2 показано "третичное сгибание" полипептидных геликсовых спиралей в пространстве. Отдельные части альфа-геликсовых спиралей полипептидных цепей отмечают латинскими буквами от А до Н (рис. 2 и рис. 3)
Все четыре третично изогнутые альфа- и бета-полипептидные цепи располагаются пространственно в определенном соотношении ("кватернерная структура"), что показано схематически на рис. 4. Они связаны между собой не настоящими химическими связями, а межмолекулярными силами.
Четыре гема гемоглобиновой молекулы расположены в форме дисков меду складками четырех альфа-, соответственно бета-полипептидных цепей (рис. 3), причем каждый гем связан с одной полипептидной цепью посредством координационной связи между Fe++-атомом гема и гистидиновым остатком полипептидной цепи (рис. 5).
Комплекс, составленный из одного гема и одной альфа-, респ. бета-полипептидной цепи, называется Сведберговой единицей. Очевидно гемоглобиновая молекула состоит из четырех Сведберговых единиц. В настоящее время принято считать, что молекулярный вес гемоглобина равен 64458, т.е. на один атом железа, соответственно приблизительно на Сведбергову единицу полагается по 16115.
Вопрос 2 Цикл трикарбоновых кислот
Цикл трикарбоновых кислот впервые был открыт английским биохимиком Кребсом. Он первым постулировал значение данного цикла для полного сгорания пирувата, главным источником которого является гликолитическое превращение углеводов. В дальнейшем было показано, что цикл трикарбоновых кислот является "фокусом", в котором сходятся практически все метаболические пути.

Итак, образовавшийся в результате окислительного декарбоксилирования пирувата ацетил-КоА вступает в цикл Кребса. Данный цикл состоит из восьми последовательных реакций (рис. 91). Начинается цикл с конденсации ацетил-КоА с оксалоацетатом и образования лимонной кислоты. (Как будет видно ниже, в цикле окислению подвергается собственно не ацетил-КоА, а более сложное соединение - лимонная кислота (трикарбоновая кислота).)
Затем лимонная кислота (шестиуглеродное соединение) путем ряда дегидрирований (отнятие водорода) и дскарбоксилирований (отщепление СО2) теряет два углеродных атома и снова в цикле Кребса появляется оксалоацетат (четырехуглеродное соединение), т. е. в результате полного оборота цикла молекула ацетил-КоА сгорает до СО2и Н2О, а молекула оксалоацетата регенерируется. Ниже приводятся все восемь последовательных реакций (этапов) цикла Кребса.
В первой реакции, катализируемой ферментом цитратсинтазой, ацетил-КоА конденсируется с оксалоацетатом. В результате образуется лимонная кислота:
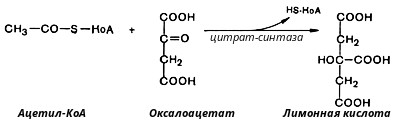
По-видимому, в данной реакции в качестве промежуточного продукта образуется связанный с ферментом цитрил-КоА. Затем последний самопроизвольно и необратимо гидролизуется с образованием цитрата и HS-KoA.
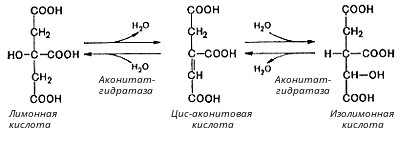
Во второй реакции цикла образовавшаяся лимонная кислота подвергается дегидратированию с образованием цис-аконитовой кислоты, которая, присоединяя молекулу воды, переходит в изолимонную кислоту. Катализирует эти обратимые реакции гидратации-дегидратации фермент аконитат-гидратаза:
В третьей реакции, которая, по-видимому, лимитирует скорость цикла Кребса, изолимонная кислота дегидрируется в присутствии НАД-зависимой изоцитратдегидрогеназы:
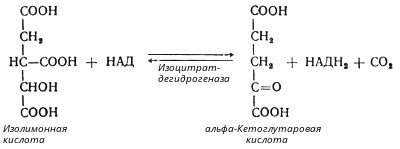
(В тканях существует два типа изоцитратдегидрогеназ: НАД- и НАДФ-зависимые. Установлено, что роль основного катализатора окисления изолимонной кислоты в цикле Кребса выполняет НАД-зависимая изоцитратдегидрогеназа.)
В ходе изоцитратдегидрогеназной реакции изолимонная кислота декарбоксилируется. НАД-зависимая изоцитратдегидрогеназа является аллостерическим ферментом, которому в качестве специфического активатора необходим АДФ. Кроме того, фермент для проявления своей активности нуждается в ионах Mg2+ или Мn2+.
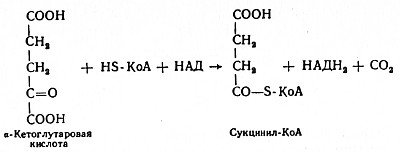
В четвертой реакции происходит окислительное декарбоксилирование α-кетоглутаровой кислоты до сукцинил-КоА. Механизм этой реакции сходен с реакцией окислительного декарбоксилирования пирувата до ацетил-КоА. α-Кетоглутаратдегидрогеназный комплекс напоминает по своей структуре пируватдегидрогеназный комплекс. Как в одном, так и в другом случае в ходе реакции принимают участие пять коферментов: TДФ, амид липоевой кислоты, HS-KoA, ФАД и НАД. Суммарно данную реакцию можно написать так:
Пятая реакция катализируется ферментом сукцинил-КоА-синтетазой. В ходе этой реакции сукцинил-КоА при участии ГДФ и неорганического фосфата превращается в янтарную кислоту (сукцинат). Одновременно происходит образование высокоэргической фосфатной связи ГТФ1 за счет высокоэргической тиоэфирной связи сукцинил-КоА:
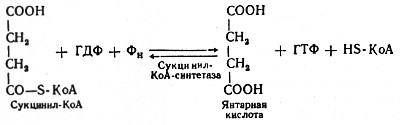
(Образовавшийся ГТФ отдает затем свою концевую фосфатную группу на АДФ, вследствие чего образуется АТФ. Образование высокоэргического нуклеозидтрифосфата в ходе сукцинил-КоА-синтетазной реакции - пример фосфорилирования на уровне субстрата.)
В шестой реакции сукцинат дегидрируется в фумаровую кислоту. Окисление сукцината катализируется сукцинатдегидрогеназой, в молекуле которой с белком ковалентно связан кофермент ФАД:
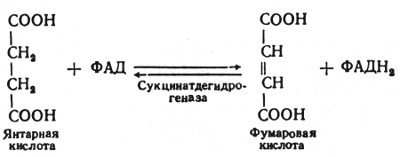
В седьмой реакции образовавшаяся фумаровая кислота гидратируется под влиянием фермента фумаратгидратазы. Продуктом данной реакции является яблочная кислота (малат). Следует отметить, что фумаратгидратаза обладает стереоспецифичностью, - в ходе данной реакции образуется L-яблочная кислота:
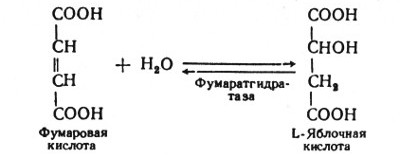
Наконец, в восьмой реакции цикла трикарбоновых кислот под влиянием митохондриальной НАД-зависимой малатдегидрогеназы происходит окисление L-малата в оксалоацетат:
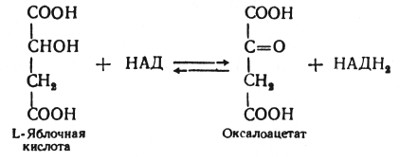
Как видно, за один оборот цикла, состоящего из восьми ферментативных реакций, происходит полное окисление ("сгорание") одной молекулы ацетил-КоА. Для непрерывной работы цикла необходимо постоянное поступление в систему ацетил-КоА, а коферменты (НАД и ФАД), перешедшие в восстановленное состояние, должны снова и снова окисляться. Это окисление осуществляется в системе переносчиков электронов (или в цепи дыхательных ферментов), локализованной в митохондриях.
Освобождающаяся в результате окисления ацетил-КоА энергия в значительной мере сосредоточивается в макроэргических фосфатных связях АТФ. Из четырех пар атомов водорода три пары переносятся через НАД на систему транспорта электронов; при этом в расчете на каждую пару в системе биологического окисления образуются три молекулы АТФ (в процессе сопряженного окислительного фосфорилирования), а всего, следовательно, девять молекул АТФ. Одна пара атомов попадает в систему транспорта электронов через ФАД, - в результате образуются 2 молекулы АТФ. В ходе реакций цикла Кребса синтезируется также 1 молекула ГТФ, что равносильно 1 молекуле АТФ. Итак, при окислении ацетил-КоА в цикле Кребса образуется 12 молекул АТФ.
Если же подсчитать энергетический эффект гликолитического расщепления глюкозы и последующего окисления пирувата до СO2 и Н2O, то он окажется значительно большим.
Как уже отмечалось, 1 молекула НАДН2 (3 молекулы АТФ) образуется при окислительном декарбоксилирова-нии пирувата в ацетил-КоА. Так как при расщеплении одной молекулы глюкозы образуются две молекулы пирувата, то при окислении их до 2 молекул ацетил-КоА и последующих двух оборотов цикла трикарбоновых кислот синтезируется 30 молекул АТФ (следовательно, окисление одной молекулы пирувата до СО2 и Н2O дает 15 молекул АТФ).
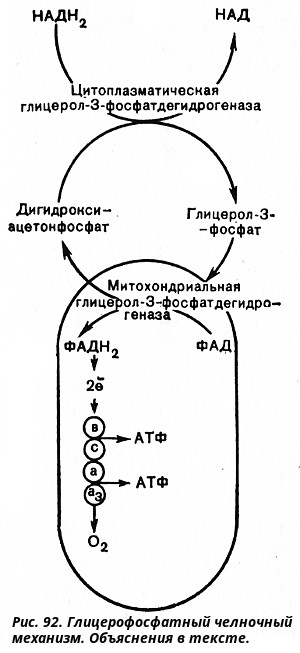
К этому надо добавить 2 молекулы АТФ, образующиеся при аэробном гликолизе, и 4 молекулы АТФ, синтезирующихся за счет окисления 2 молекул внемитохондриального НАДН2, которые образуются при окислении 2 молекул глицеральдегид-3-фосфата в дегидрогеназной реакции. Итого получим, что при расщеплении в тканях 1 молекулы глюкозы по уравнению: C6H1206 + 602 -> 6СO2 + 6Н2O синтезируется 36 молекул АТФ, что способствует накоплению в макроэргических фосфатных связях аденозинтрифосфата 36 X 34,5 ~ 1240 кДж (или, по другим данным, 36 Х 38 ~ 1430 кДж) свободной энергии. Другими словами, из всей освобождающейся при аэробном окислении глюкозы свободной энергии (окодо 2840 кДж) до 50% ее аккумулируется в митохондриях в форме, которая может быть использована для выполнения различных физиологических функций. Несомненно, что в энергетическом отношении полное расщепление глюкозы является более эффективным процессом, чем гликолиз. Необходимо отметить, что образовавшиеся в процессе превращения глицеральдегид-3-фосфата 2 молекулы НАДН2 в дальнейшем при окислении дают не 6 молекул АТФ, а только 4. Дело в том, что сами молекулы внемитохондриального НАДН2 не способны проникать через мембрану внутрь митохондрий. Однако отдаваемые ими электроны могут включаться в митохондриальную цепь биологического окисления с помощью так называемого глицерофосфатного челночного механизма (рис. 92). Как видно на рисунке, цитоплазматический НАДН2 сначала реагирует с цитоплазматическим дигидроксиацетонфосфатом, образуя глицерол-3-фосфат. Реакция катализируется НАД-зависимой цитоплазматической глицерол-3-фосфат-дегидрогеназой:
Дигидрооксиацетонфосфат + НАДН2 <---> глицерол-3-фосфат + НАД
Образовавшийся глицерол-3-фосфат легко проникает через митохондриальную мембрану. Внутри митохондрии другая (митохондриальная) глицерол-3-фосфат-дегидрогеназа (флавиновый фермент) снова окисляет глицерол-3-фосфат до дигидроксиацетонфосфата:
Глицерол-З-фосфат + ФАД <---> Дигидроксиацетонфосфат + фАДН2
Восстановленный флавопротеид (фермент - ФАДН2) вводит, на уровне KoQ приобретенные им электроны в цепь биологического окисления и сопряженного с ним окислительного фосфорилирования, а дигидроксиацетонфосфат выходит из митохондрий в цитоплазму и может вновь взаимодействовать с цитоплазматическим НАДН2. Таким образом, пара электронов (из одной молекулы цитоплазматического НАДН2), вводимая в дыхательную цепь с помощью глицерофосфатного челночного механизма, дает не 3 АТФ, а 2 АТФ.
В настоящее время четко установлено, что глицерофосфатный челночный механизм имеет место в клетках печени. Относительно других тканей этот вопрос пока не выяснен.
Если же не считать энергии, затрачиваемой на проникновение НАДН2 в митохондрии, то в результате полного окисления 1 молекулы глюкозы может образоваться 38 молекул АТФ.
Цикл трикарбоновых кислот (ЦТК ) или цикл лимонной кислоты или цикл Кребса – путь окислительных превращений ди- и трикарбоновых кислот, образующихся в качестве промежуточных продуктов при распаде и синтезе белков, жиров и углеводов.
Цикл трикарбоновых кислот представлен в клетках всех организмов: растений, животных и микроорганизмов.
Этот цикл является основой метаболизма и выполняет две важных функции:
- снабжения организма энергией;
- интеграции всех главных метаболических потоков, как катаболических (биорасщепление), так и анаболических (биосинтез).
Напомню, что реакции аэробного гликолиза локализованы в цитоплазме клетки и приводят к образованию пирувата (ПВК ).
Последующие превращения пирувата протекают в матриксе митохондрий.
В матриксе пируват превращается в ацетил-КоА – макроэргическое соединение. Реакция катализируется ферментом НАД-зависимой пируватдекарбоксилазой:

Восстановленная форма НАДН∙Н+, образовавшаяся в результате этой реакции, поступает в дыхательную цепь и генерирует 6 молекул АТФ (в пересчете на 1 молекулу глюкозы).
ЦТК представляет собой последовательность из восьми реакций, протекающих в матриксемитохондрий (Рис. 1):

Рис. 1. Схема цикла трикарбоновых кислот
1) Необратимая реакция конденсации ацетил-КоА со щавелевоуксусной кислотой (оксалоацетатом), катализируемая ферментом цитратсинтетазой, с образованием лимонной кислоты (цитрата ).
2) Обратимая реакция изомеризация лимонной кислоты (цитрата ) в изолимонную кислоту(изоцитрат ), в процессе которой происходит перенос гидроксигруппы к другому атому углерода, катализируется ферментом аконитазой.
Реакция идёт через образование промежуточного продукта
цис-аканитовой кислоты (цис-аконитата ).
3) Необратимая реакция окислительного декарбоксилирования изолимонной кислоты (изоцитрата): гидроксигруппа изолимонной кислоты окисляется до карбонильной группы с помощью окисленной формы НАД+ и одновременно отщепляется карбоксильная группа в
β-положении с образованием α-кетоглутаровой кислоты (α-кетоглутарата ). Промежуточный продукт этой реакции щавелевоянтарная кислота (оксалосукцинат ).
Это первая реакция цикла, в которой происходит восстановление окисленной формы НАД+-кофермента до НАДН∙Н+, фермента изоцитратдегидрогеназы.
Восстановленная форма НАДН∙Н поступает в дыхательную цепь, там окисляется до НАД+, что приводит к образованию 2 молекул АТФ.
4) Обратимая реакция окислительного декарбоксилирования
α-кетоглутаровой кислоты до макроэргического соединения сукцинил-КоА. Реакцию катализирует фермент 2-оксоглутаратдегидрогеназный комплекс.
5) Реакция является единственной в цикле реакцией субстратного фосфорилирования; катализируется ферментом сукцинил-КоА-синтетазой. В этой реакции сукцинил-КоА при участиигуанодиндифосфата (ГДФ ) и неорганического фосфата (H 3 PO 4 ) превращается в янтарную кислоту (сукцинат ).
Одновременно происходит синтез макроэргического соединения ГТФ за счёт макроэргической связи тиоэфирной связи сукцинил-КоА.
6) Реакция дегидрирования янтарной кислоты (сукцината ) с образованием фумаровой кислоты(фумарата ).
Реакция катализируется сложным ферментом сукцинатдегидрогеназой, в молекуле которой кофермент ФАД+ ковалентно связан, а белковой частью фермента. Окисленная форма ФАД+ в результате реакции восстанавливается до ФАД∙Н2.
Восстановленная форма ФАД∙Н2 поступает в дыхательную цепь, там регенерирует до окисленной формы ФАД+, что приводит к образованию двух молекул АТФ.
7) Реакция гидратации фумаровой кислоты (фумарата ) до яблочной кислоты (малата ). Реакция катализируется ферментом фумаразой.
8) Реакция дегидрирования яблочной кислоты до щавелеуксусной кислоты (оксалоацетата ). Реакция катализируется ферментом НАД+-зависимой-малатдегидрогеназой.
В результате реакции окисленная форма НАД восстанавливается до восстановленной формы НАДН∙Н+.
Восстановленная форма НАДН∙Н поступает в дыхательную цепь, там окисляется до НАД+, что приводит к образованию 2 молекул АТФ.
Суммарное уравнение ЦТК можно записать следующим образом:
Ацетил-КоА + 3НАД+ + ФАД+ + ГДФ + H 3 PO 4 =
2 CO 2 + H 2 O + HS -КоА + 3НАДН∙Н + ФАД∙Н2 + ГТФ
Как видно из схемы суммарного уравнения ЦТК в этом процессе восстанавливаются:
- три молекулы НАДН∙Н (реакции 3, 4, 8);
- одна молекула ФАД∙Н2 (реакция 6).
При аэробном окислении из этих молекул в электрон-транспортной цепи в процессе окислительного фосфорилирования образуется при окислении:
- одной молекулы НАДН∙Н – 3 молекулы АТФ;
- одной молекулы ФАД∙Н2 – 2 молекулы АТФ.
- одна молекула ГТФ образуется в реакции субстратного фосфорилирования (реакция 5 ).
Всё это составит: 9 (3х3) АТФ + 2 АТФ + 1 АТФ (ГТФ ) = 12 АТФ. Следовательно, энергетический баланс окисления ацетил-КоА (2 молекулы пирувата из аэробного гликолиза ) в ЦТК составляет 24 молекулы АТФ.
Полное окисление глюкозы:
8 молекул АТФ гликолиза + 6 молекул АТФ окислительного декарбоксилирование пирувата вцетил-КоА + 24 молекулы АТФ ЦТК =
38 молекул АТФ на молекулу глюкозы.