
Вопрос 1 Общий план строения клетки.
Все клеточные формы жизни на Земле можно разделить на два надцарства на основании строения составляющих их клеток:
- прокариоты (доядерные) — более простые по строению и возникли в процессе эволюции раньше;
- эукариоты (ядерные) — более сложные, возникли позже. Клетки, составляющие тело человека, являются эукариотическими.
Несмотря на многообразие форм, организация клеток всех живых организмов подчинена единым структурным принципам.
Содержимое клетки отделено от окружающей среды плазматической мембраной, или плазмалеммой. Внутри клетка заполнена цитоплазмой, в которой расположены различные органоиды и клеточные включения, а также генетический материал в виде молекулы ДНК. Каждый из органоидов клетки выполняет свою особую функцию, а в совокупности все они определяют жизнедеятельность клетки в целом.
Прокариотическая клетка[править | править вики-текст]
Прокариоты (от лат. pro — перед, до и греч. κάρῠον — ядро, орех) — организмы, не обладающие, в отличие от эукариот, оформленным клеточным ядром и другими внутренними мембранными органоидами (за исключением плоских цистерн у фотосинтезирующих видов, например, у цианобактерий). Единственная крупная кольцевая (у некоторых видов — линейная) двухцепочечная молекула ДНК, в которой содержится основная часть генетического материала клетки (так называемыйнуклеоид) не образует комплекса с белками-гистонами (так называемого хроматина). К прокариотам относятся бактерии, в том числе цианобактерии (сине-зелёные водоросли), и археи. Потомками прокариотических клеток являются органеллы эукариотических клеток — митохондрии и пластиды. Основное содержимое клетки, заполняющее весь её объём, — вязкая зернистая цитоплазма.
Эукариотическая клетка[править | править вики-текст]
Эукариоты (эвкариоты) (от греч. ευ — хорошо, полностью и κάρῠον — ядро, орех) — организмы, обладающие, в отличие от прокариот, оформленным клеточнымядром, отграниченным от цитоплазмы ядерной оболочкой. Генетический материал заключён в нескольких линейных двухцепочных молекулах ДНК (в зависимости от вида организмов их число на ядро может колебаться от двух до нескольких сотен), прикреплённых изнутри к мембране клеточного ядра и образующих у подавляющего большинства (кроме динофлагеллят) комплекс с белками-гистонами, называемый хроматином. В клетках эукариот имеется система внутренних мембран, образующих, помимо ядра, ряд других органоидов (эндоплазматическая сеть, аппарат Гольджи и др.). Кроме того, у подавляющего большинства имеются постоянные внутриклеточные симбионты-прокариоты — митохондрии, а у водорослей и растений — также и пластиды.
Строение прокариотической клетки[править | править вики-текст]



Клетки двух основных групп прокариот — бактерий и архей — похожи по структуре, характерными их признаками являются отсутствие ядра и мембранных органелл.
Основными компонентами прокариотической клетки являются:
- Клеточная стенка, которая окружает клетку извне, защищает её, придаёт устойчивую форму, предотвращающую от осмотическогоразрушения. У бактерий клеточная стенка состоит из пептидогликана (муреина), построенного из длинных полисахаридных цепей, соединенных между собой короткими пептидными перемычками. По строению клеточной стенки различают две группы бактерий:
- Грамположительные бактерии (например роды Staphylococcus, Bacillus, Lactobacillus[4]) — имеют более простую структуру клеточной стенки, состоящую почти исключительно из муреина;
- Грамотрицательные бактерии (например роды Salmonella, Escherichia, Azotobacter[4]) — клеточная стенка содержит меньше пептидогликана и имеет дополнительную внешнюю мембрану, которая состоит из фосфолипидов.
Клеточная стенка архей не содержит муреина, а построена в основном из разнообразных белков и полисахаридов[3].
- Капсула — имеющаяся у некоторых бактерий слизистая оболочка, расположенная снаружи от клеточной стенки. Состоит в основном из разнообразных белков, углеводов и уроновых кислот. Капсулы защищают клетки от высыхания, могут помогать бактериям вколониях удерживаться вместе, а индивидуальным бактериям — прикрепляться к различным субстратам. Кроме этого, капсулы предоставляют клетке дополнительную защиту: например, капсулированные штаммы пневмококков свободно размножаются в организме и вызывают воспаление легких, тогда как некапсулированные быстро уничтожаются иммунной системой и являются абсолютно безвредными[4].
- Пили или ворсинки — тонкие волоскоподобные выросты, что присутствуют на поверхности бактериальных клеток. Существуют различные типы пилей, из которых наиболее распространенными являются:
- Фимбрии — пили, которые служат для прикрепления. Например, возбудитель гонореи — Neisseria gonorrhoeae использует фимбрии для удержания на слизистой оболочке хозяина.
- Половые пили (F-пили) — задействованы в процессе конъюгации у бактерий[3].
- Жгутики — органеллы движения некоторых бактерий. Бактериальный жгутик построен значительно проще эукариотического, и он в 10 раз тоньше, внешне не покрыт плазматической мембраной и состоит из одинаковых молекул белков, которые образуют цилиндр. В мембране жгутик закреплен при помощи базального тела[3].
- Плазматическая и внутренние мембраны. Общий принцип устройства клеточных мембран не отличается от эукариот, однако химическом составе мембраны есть немало различий, в частности, в мембранах прокариот отсутствуют молекулы холестерина и некоторых липидов, присущих мембранам эукариот. Большинство прокариотических клеток (в отличие от эукариотических) не имеют внутренних мембран, которые разделяют цитоплазму на отдельные компартменты. Только у некоторых фотосинтетических и аэробных бактерий плазмалемма образует вгибание внутрь клетки, что выполняет соответствующие метаболические функции[3].
- Нуклеоид — не ограниченный мембранами участок цитоплазмы, в котором расположена кольцевая молекула ДНК — «бактериальная хромосома», где хранится весь генетический материал клетки[3].
- Плазмиды — небольшие дополнительные кольцевые молекулы ДНК, несущие обычно всего несколько генов. Плазмиды, в отличие от бактериальной хромосомы, не являются обязательным компонентом клетки. Обычно они придают бактерии определенные полезные для неё свойства, такие как устойчивость к антибиотикам, способность усваивать из среды определенные энергетические субстраты, способность инициировать половой процесс и т. д.[4][3]
- Рибосомы прокариот, как и у всех других живых организмов, отвечают за осуществление процесса трансляции (одного из этапов биосинтеза белка). Однако бактериальные рибосомы несколько меньше, чем эукариотические (коэффициенты седиментации 70S и 80S соответственно), и имеют другой состав белков и РНК. Из-за этого бактерии, в отличие от эукариот, чувствительны к таким антибиотикам, как эритромицин и тетрациклин, которые избирательно действуют на 70S-рибосомы[4].
- Эндоспоры — окруженные плотной оболочкой структуры, содержащие ДНК бактерии и обеспечивающее выживание в неблагоприятных условиях. К образованию эндоспор способны лишь некоторые виды прокариот, например представители родов Clostridium (C. tetani — возбудитель столбняка, C. botulinum — возбудительботулизма, C. perfringens — возбудитель газовой гангрены и т. п.) и Bacillus (в частности B. anthracis — возбудитель сибирской язвы). Для образования эндоспоры клетка реплицирует свою ДНК и окружает копию плотной оболочкой, из созданной структуры удаляется избыток воды, и в ней замедляется метаболизм[3]. Споры бактерий могут выдерживать довольно жесткие условия среды, такие как длительное высушивание, кипячение, коротковолновое облучение и др.[4]
Строение эукариотической клетки[править | править вики-текст]

Поверхностный комплекс животной клетки[править | править вики-текст]
Состоит из гликокаликса, плазмалеммы и расположенного под ней кортикального слоя цитоплазмы. Плазматическая мембрана называется также плазмалеммой, наружной клеточной мембраной. Это биологическая мембрана, толщиной около 10 нанометров. Обеспечивает в первую очередь разграничительную функцию по отношению к внешней для клетки среде. Кроме этого она выполняет транспортную функцию. На сохранение целостности своей мембраны клетка не тратит энергии: молекулы удерживаются по тому же принципу, по которому удерживаются вместе молекулы жира — гидрофобным частям молекул термодинамически выгоднее располагаться в непосредственной близости друг к другу. Гликокаликс представляет собой «заякоренные» в плазмалемме молекулы олигосахаридов, полисахаридов,гликопротеинов и гликолипидов. Гликокаликс выполняет рецепторную и маркерную функции. Плазматическая мембрана животных клеток в основном состоит изфосфолипидов и липопротеидов со вкрапленными в неё молекулами белков, в частности, поверхностных антигенов и рецепторов. В кортикальном (прилегающем к плазматической мембране) слое цитоплазмы находятся специфические элементы цитоскелета — упорядоченные определённым образом актиновые микрофиламенты. Основной и самой важной функцией кортикального слоя (кортекса) являются псевдоподиальные реакции: выбрасывание, прикрепление и сокращение псевдоподий. При этом микрофиламенты перестраиваются, удлиняются или укорачиваются. От структуры цитоскелета кортикального слоя зависит также форма клетки (например, наличие микроворсинок).
Структура цитоплазмы[править | править вики-текст]
Жидкую составляющую цитоплазмы также называют цитозолем. Под световым микроскопом казалось, что клетка заполнена чем-то вроде жидкой плазмы или золя, в котором «плавают» ядро и другие органоиды. На самом деле это не так. Внутреннее пространство эукариотической клетки строго упорядочено. Передвижение органоидов координируется при помощи специализированных транспортных систем, так называемых микротрубочек, служащих внутриклеточными «дорогами», и специальных белков динеинов и кинезинов, играющих роль «двигателей». Отдельные белковые молекулы также не диффундируют свободно по всему внутриклеточному пространству, а направляются в необходимые компартменты при помощи специальных сигналов на их поверхности, узнаваемых транспортными системами клетки.

Эндоплазматический ретикулум[править | править вики-текст]
В эукариотической клетке существует система переходящих друг в друга мембранных отсеков (трубок и цистерн), которая называется эндоплазматическим ретикулумом (или эндоплазматическая сеть, ЭПР или ЭПС). Ту часть ЭПР, к мембранам которого прикреплены рибосомы, относят к гранулярному (или шероховатому) эндоплазматическому ретикулуму, на его мембранах происходит синтез белков. Те компартменты, на стенках которых нет рибосом, относят к агранулярному (или гладкому) ЭПР, принимающему участие в синтезе липидов. Внутренние пространства гладкого и гранулярного ЭПР не изолированы, а переходят друг в друга и сообщаются с просветом ядерной оболочки.
Аппарат Гольджи[править | править вики-текст]
Аппарат Гольджи представляет собой стопку плоских мембранных цистерн, несколько расширенных ближе к краям. В цистернах аппарата Гольджи созревают некоторые белки, синтезированные на мембранах гранулярного ЭПР и предназначенные для секреции или образования лизосом. Аппарат Гольджи асимметричен — цистерны располагающиеся ближе к ядру клетки (цис-Гольджи) содержат наименее зрелые белки, к этим цистернам непрерывно присоединяются мембранные пузырьки — везикулы, отпочковывающиеся от эндоплазматического ретикулума. По-видимому, при помощи таких же пузырьков происходит дальнейшее перемещение созревающих белков от одной цистерны к другой. В конце концов от противоположного конца органеллы (транс-Гольджи) отпочковываются пузырьки, содержащие полностью зрелые белки.
Ядро[править | править вики-текст]
Клеточное ядро содержит молекулы ДНК, на которых записана генетическая информация организма. В ядре происходит репликация — удвоение молекул ДНК, а также транскрипция — синтез молекул РНК на матрице ДНК. В ядре же синтезированные молекулы РНК претерпевают некоторые модификации (например, в процессе сплайсинга из молекул матричной РНК исключаются незначащие, бессмысленные участки), после чего выходят в цитоплазму. Сборка рибосом также происходит в ядре, в специальных образованиях, называемыхядрышками. Компартмент для ядра — кариотека — образован за счёт расширения и слияния друг с другом цистерн эндоплазматической сети таким образом, что у ядра образовались двойные стенки за счёт окружающих его узких компартментов ядерной оболочки. Полость ядерной оболочки называется люменом илиперинуклеарным пространством. Внутренняя поверхность ядерной оболочки подстилается ядерной ламиной, жесткой белковой структурой, образованной белками-ламинами, к которой прикреплены нити хромосомной ДНК. В некоторых местах внутренняя и внешняя мембраны ядерной оболочки сливаются и образуют так называемые ядерные поры, через которые происходит материальный обмен между ядром и цитоплазмой.
Лизосомы[править | править вики-текст]
Лизосома — небольшое тельце, ограниченное от цитоплазмы одинарной мембраной. В ней находятся литические ферменты, способные расщепить все биополимеры. Основная функция — автолиз — то есть расщепление отдельных органоидов, участков цитоплазмы клетки.
Цитоскелет[править | править вики-текст]
К элементам цитоскелета относят белковые фибриллярные структуры, расположенные в цитоплазме клетки: микротрубочки, актиновые и промежуточные филаменты. Микротрубочки принимают участие в транспорте органелл, входят в состав жгутиков, из микротрубочек строится митотическое веретено деления. Актиновые филаменты необходимы для поддержания формы клетки, псевдоподиальных реакций. Роль промежуточных филаментов, по-видимому, также заключается в поддержании структуры клетки. Белки цитоскелета составляют несколько десятков процентов от массы клеточного белка.
Центриоли[править | править вики-текст]
Центриоли представляют собой цилиндрические белковые структуры, расположенные вблизи ядра клеток животных (у растений центриолей нет, за исключением низших водорослей). Центриоль представляет собой цилиндр, боковая поверхность которого образована девятью наборами микротрубочек. Количество микротрубочек в наборе может колебаться для разных организмов от 1 до 3.
Вокруг центриолей находится так называемый центр организации цитоскелета, район в котором группируются минус концы микротрубочек клетки.
Перед делением клетка содержит две центриоли, расположенные под прямым углом друг к другу. В ходе митоза они расходятся к разным концам клетки, формируя полюса веретена деления. После цитокинеза каждая дочерняя клетка получает по одной центриоли, которая удваивается к следующему делению. Удвоение центриолей происходит не делением, а путём синтеза новой структуры, перпендикулярной существующей.
Центриоли, по-видимому, гомологичны базальным телам жгутиков и ресничек.
Митохондрии[править | править вики-текст]
Митохондрии — особые органеллы клетки, основной функцией которых является синтез АТФ — универсального носителя энергии. Дыхание (поглощение кислорода и выделение углекислого газа) происходит также за счёт энзиматических систем митохондрий.
Внутренний просвет митохондрий, называемый матриксом, отграничен от цитоплазмы двумя мембранами, наружной и внутренней, между которыми располагаетсямежмембранное пространство. Внутренняя мембрана митохондрии образует складки, так называемые кристы. В матриксе содержатся различные ферменты, принимающие участие в дыхании и синтезе АТФ. Центральное значение для синтеза АТФ имеет водородный потенциал внутренней мембраны митохондрии.
Митохондрии имеют свой собственный ДНК-геном и прокариотические рибосомы, что, безусловно, указывает на симбиотическое происхождение этих органелл. В ДНК митохондрий закодированы совсем не все митохондриальные белки, большая часть генов митохондриальных белков находятся в ядерном геноме, а соответствующие им продукты синтезируются в цитоплазме, а затем транспортируются в митохондрии. Геномы митохондрий отличаются по размерам: например геном человеческих митохондрий содержит всего 13 генов. Самое большое число митохондриальных генов (97) из изученных организмов имеет простейшее Reclinomonas americana.
Сравнение прокариотической и эукариотической клеток[править | править вики-текст]
Наиболее важным отличием эукариот от прокариот долгое время считалось наличие оформленного ядра и мембранных органоидов. Однако к 1970—1980-м гг. стало ясно, что это лишь следствие более глубинных различий в организации цитоскелета. Некоторое время считалось, что цитоскелет свойственен только эукариотам, но в середине 1990-х гг. белки, гомологичные основным белкам цитоскелета эукариот, были обнаружены и у бактерий.

Именно наличие специфическим образом устроенного цитоскелета позволяет эукариотам создать систему подвижных внутренних мембранных органоидов. Кроме того, цитоскелет позволяет осуществлять эндо- и экзоцитоз (как предполагается, именно благодаря эндоцитозу в эукариотных клетках появились внутриклеточные симбионты, в том числе митохондрии и пластиды). Другая важнейшая функция цитоскелета эукариот — обеспечение деления ядра (митоз и мейоз) и тела (цитотомия) эукариотной клетки (деление прокариотических клеток организовано проще). Различия в строении цитоскелета объясняют и другие отличия про- и эукариот — например, постоянство и простоту форм прокариотических клеток и значительное разнообразие формы и способность к её изменению у эукариотических, а также относительно большие размеры последних. Так, размеры прокариотических клеток составляют в среднем 0,5—5 мкм, размеры эукариотических — в среднем от 10 до 50 мкм. Кроме того, только среди эукариот попадаются поистине гигантские клетки, такие как массивные яйцеклетки акул или страусов (в птичьем яйце весь желток — это одна огромная яйцеклетка), нейроны крупных млекопитающих, отростки которых, укрепленные цитоскелетом, могут достигать десятков сантиметров в длину.
|
Сравнительная характеристика клеток эукариот и прокариот[4]
|
||
| Признак | Прокариоты | Эукариоты |
| Размеры клеток | Средний диаметр 0,5—10 мкм | Средний диаметр 10—100 мкм |
|
Организация генетического материала
|
||
| Форма, количество и расположение молекулДНК | Обычно имеется одна кольцевая молекула ДНК, размещенная в цитоплазме | Обычно есть несколько линейных молекул ДНК — хромосом, локализованных вядре |
| Компактизация ДНК | У бактерий ДНК компактизируется без участия гистонов[5]. Уархей ДНК ассоциирована с белками гистонами[6] | Имеется хроматин: ДНК компактизируется в комплексе с белками гистонами[5]. |
| Организация генома | У бактерий экономный геном: отсутствуют интроны и большие некодирующие участки[7]. Гены объединены вопероны[5]. У архей имеются интронные участки особой структуры[8]. |
Большей частью геном не экономный: имеется экзон-интронная организация генов, большие участки некодирующей ДНК[7] Гены не объединены в опероны[5]. |
|
Деление
|
||
| Тип деления | Простое бинарное деление | Мейоз или митоз |
| Образование веретена деления | Веретено деления не образуется | Веретено деления образуется |
|
Органеллы
|
||
| Тип рибосом | 70S рибосомы | 80S рибосомы |
| Наличие мембранных органелл | Окруженные мембранами органеллы отсутствуют, иногда плазмалемма образует выпячивание внутрь клетки | Имеется большое количество одномембранных и двумембранных органелл |
| Тип жгутика | Жгутик простой, не содержит микротрубочки, не окружен мембраной, диаметр около 20 нм | Жгутики состоят из микротрубочек, расположенных по принципу «9+2», окружены плазматической мембраной, диаметр около 200 нм |
Вопрос 2 Биосинтез белка.
Пластический обмен (ассимиляция или анаболизм) – совокупность реакций биологического синтеза. Название этого вида обмена отражает его сущность: из веществ, поступающих в клетку из вне, образуются вещества, подобные веществам клетки.
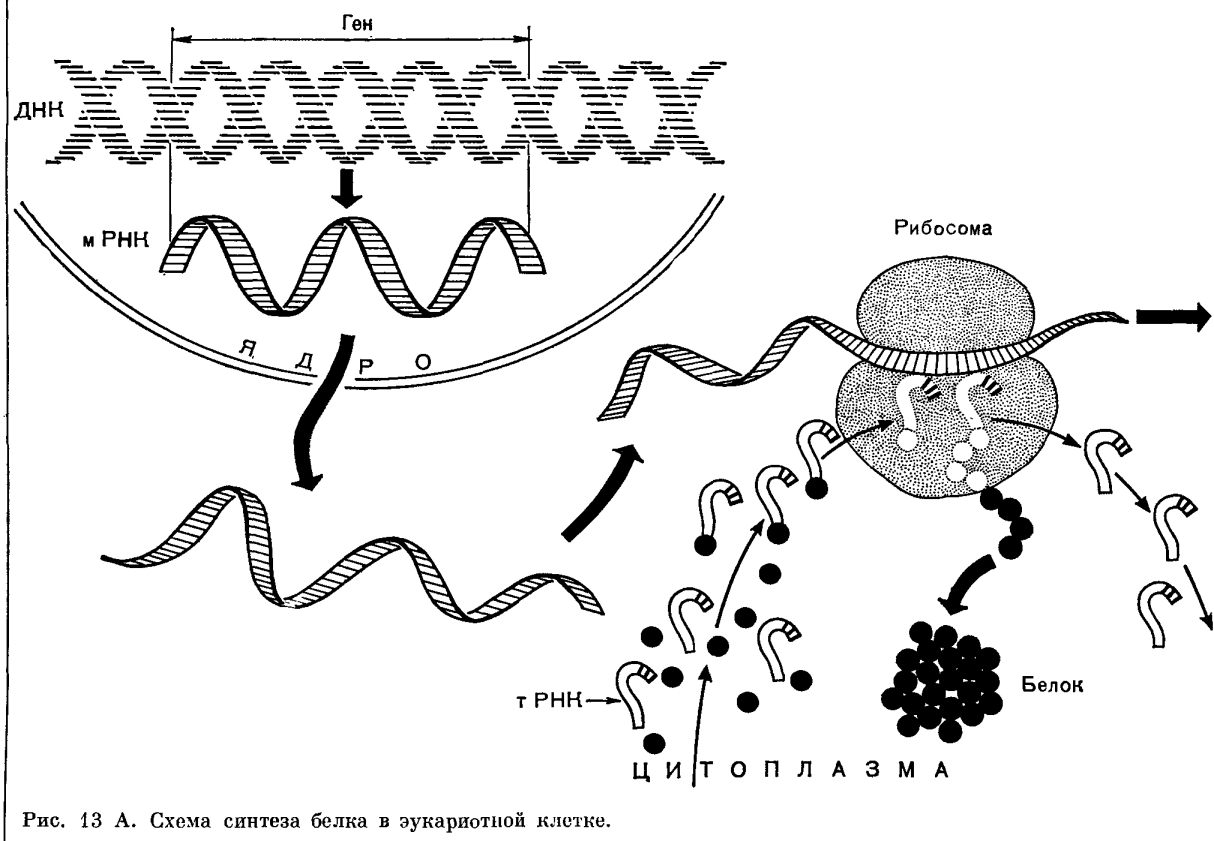
Рассмотрим одну из важнейших форм пластического обмена – биосинтез белков. Биосинтез белков осуществляется во всех клетках про -и эукариот. Информация о первичной структуре (порядке аминокислот) белковой молекулы закодирована последовательностью нуклеотидов в соответствующем участке молекулы ДНК — гене.
Ген— это участок молекулы ДНК, определяющий порядок аминокислот в молекуле белка. Следовательно, от порядка нуклеотидов в гене зависит порядок аминокислот в полипептиде, т.е. его первичная структура, от которой в свою очередь зависят все другие структуры, свойства и функции белковой молекулы.
Система записи генетической информации в ДНК (и - РНК) в виде определенной последовательности нуклеотидов называется генетическим кодом. Т.е. единица генетического кода (кодон) — это триплет нуклеотидов в ДНК или РНК, кодирующий одну аминокислоту.
Всего генетический код включает 64 кодона, из них 61 кодирующий и 3 некодирующих (кодоны-терминаторы, свидетельствующие об окончании процесса трансляции).
Кодоны-терминаторы в и - РНК: УАА, УАГ, УГА, в ДНК: АТТ, АТЦ, АЦТ.
Начало процесса трансляции определяет кодон-инициатор (АУГ, в ДНК — ТАЦ), кодирующий аминокислоту метионин. Этот кодон первым входит в рибосому. Впоследствии метионин, если он не предусмотрен в качестве первой аминокислоты данного белка, отщепляется.
Генетический код обладает характерными свойствами.
1. Универсальность — код одинаков для всех организмов. Один и тот же триплет (кодон) в любом организме кодирует одну и ту же аминокислоту.
2. Специфичность — каждый кодон шифрует только одну аминокислоту.
3. Вырожденность — большинство аминокислот могут кодироваться несколькими кодонами. Исключение составляют 2 аминокислоты — метионин и триптофан, имеющие лишь по одному варианту кодона.
4. Между генами имеются «знаки препинания» — три специальных триплета (УАА, УАГ, УГА), каждый из которых обозначает прекращение синтеза полипептидной цепи.
5. Внутри гена «знаков препинания» нет.
Для того, чтобы синтезировался белок, информация о последовательности нуклеотидов в его первичной структуре должна быть доставлена к рибосомам. Этот процесс включает два этапа – транскрипцию и трансляцию.
Транскрипция (переписывание) информации происходит путем синтеза на одной из цепей молекулы ДНК одноцепочной молекулы РНК, последовательность нуклеотидов которой точно соответствует последовательности нуклеотидов матрицы – полинуклеотидной цепи ДНК.
Она (и - РНК) является посредником, передающим информацию от ДНК к месту сборки молекул белка в рибосоме. Синтез и - РНК (транскрипция) происходит следующим образом. Фермент (РНК - полимераза) расщепляет двойную цепочку ДНК, и на одной из ее цепей (кодирующей) по принципу комплементарности выстраиваются нуклеотиды РНК. Синтезированная таким образом (матричный синтез) молекула и - РНК выходит в цитоплазму, и на один ее конец нанизываются малые субъединицы рибосом.
Второй этап в биосинтезе белка — трансляция — это перевод последовательности нуклеотидов в молекуле и - РНК в последовательность аминокислот в полипептиде. У прокариот, не имеющих оформленного ядра, рибосомы могут связываться с вновь синтезированной молекулой и - РНК сразу же после ее отделения от ДНК или даже до полного завершения ее синтеза. У эукариот и - РНК сначала должна быть доставлена через ядерную оболочку в цитоплазму. Перенос осуществляется специальными белками, которые образуют комплекс с молекулой и - РНК. Кроме функций переноса эти белки защищают и - РНК от повреждающего действия цитоплазматических ферментов.
В цитоплазме на один из концов и - РНК (а именно на тот, с которого начинается синтез молекулы в ядре) вступает рибосома и начинается синтез полипептида. По мере продвижения по молекуле РНК рибосома транслирует триплет за триплетом, последовательно присоединяя аминокислоты к растущему концу полипептидной цепи. Точное соответствие аминокислоты коду триплета и - РНК обеспечивается т - РНК.
Транспортные РНК (т - РНК) «приносят» аминокислоты в большую субъединицу рибосомы. Молекула т - РНК имеет сложную конфигурацию. На некоторых участках ее между комплементарными нуклеотидами образуются водородные связи, и молекула по форме напоминает лист клевера. На ее верхушке расположен триплет свободных нуклеотидов (антикодон), который соответствует определенной аминокислоте, а основание служит местом прикрепления этой аминокислоты (рис. 1).

Рис. 1. Схема строения транспортной РНК: 1 — водородные связи; 2 — антикодон; 3 —место прикрепления аминокислоты.
Каждая т - РНК может переносить только свою аминокислоту. Т-РНК активируется специальными ферментами, присоединяет свою аминокислоту и транспортирует ее в рибосому. Внутри рибосомы в каждый данный момент находится всего два кодона и-РНК. Если антикодон т-РНК является комплементарным кодону и-РНК, то происходит временное присоединение т-РНК с аминокислотой к и-РНК. Ко второму кодону присоединяется вторая т-РНК, несущая свою аминокислоту. Аминокислоты располагаются рядом в большой субъединице рибосомы, и с помощью ферментов между ними устанавливается пептидная связь. Одновременно разрушается связь между первой аминокислотой и ее т-РНК, и т-РНК уходит из рибосомы за следующей аминокислотой. Рибосома перемещается на один триплет, и процесс повторяется. Так постепенно наращивается молекула полипептида, в которой аминокислоты располагаются в строгом соответствии с порядком кодирующих их триплетов (матричный синтез) (рис. 2).

Рис. 2. Схема бисинтеза белка: 1 — и-РНК; 2 - субъединицы рибосомы; 3 — т-РНК с аминокислотами; 4 — т-РНК без аминокислот; 5 — полипептид; 6 — кодон и-РНК; 7- антикодон т-РНК.
Одна рибосома способна синтезировать полную полипептидную цепь. Однако, нередко по одной молекуле и-РНК движется несколько рибосом. Такие комплексы называются полирибосомами. После завершения синтеза полипептидная цепочка отделяется от матрицы – молекулы и-РНК, сворачивается в спираль и приобретает свойственную ей (вторичную, третичную или четвертичную) структуру. Рибосомы работают очень эффективно: в течение 1с бактериальная рибосома образует полипептидную цепь из 20 аминокислот.
,
